NEWSLETTER 05/2014 20.05.2014
|
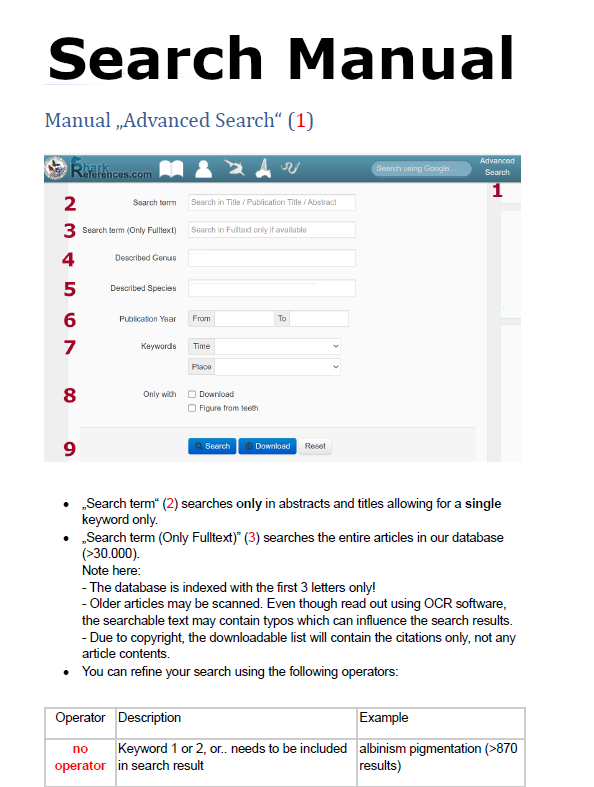
|
Pollerspöck, J. 2014, Bibliography database of living/fossil sharks, rays and chimaeras (Chondrichtyes: Elasmobranchii, Holocephali), www.shark-references.com, World Wide Web electronic publication, Version 2014 |
|
|
PARTNERS OF SHARK-REFERENCES:
New partner:
- Canary Islands Shark Alliance (Alianza por los Tiburones de Canarias) (Homepage/facebook)
- Fernando Frias Reis, Presidente Alianza por los Tiburones de Canarias (Homepage)
Would you like to become a shark-reference partner? Please contanct me per E-mail!
Partner in Google-Maps: 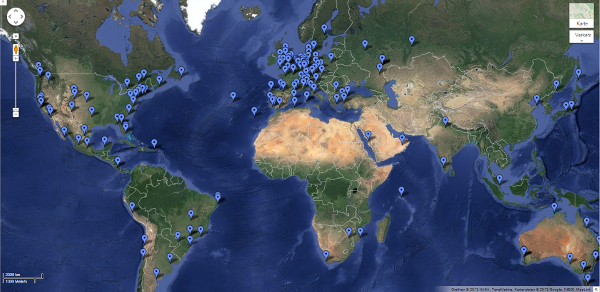
|
|
New images at shark-references:
Many thanks to the following persons for the permission to use their images:
Please support shark-references and send your images to: info@shark-references.com |
|
Missing papers:
Many thanks to all friends of shark-references, who send me some missing papers last month!
Shark-References would kindly like to ask you for your contribution to this project.
At the moment I search e.g. the following papers:
FOSSIL:
ANTUNES, M.T. & BALBINO, A.C. & FREITAS, I. 2005 Early (18th century) discovery of Cretaceous fishes from Chapada do Araripe, Ceará, Brazil - Specimens kept at the ‘Academia das Ciências de Lisboa' Museum. Comptes Rendus - Palevol, 4 (4): 375-384
BECKER, M.A. & CHAMBERLAIN, R.B. & CHAMBERLAIN, J.A. 2007 The paradox of large carcharhinoid-type shark vertebrae in the Upper Cretaceous of New Jersey. In: Contributions to the Paleontology of New Jersey (II), E Rainforth, ed., Geological Association of New Jersey. Proceedings of the XXIV Annual Meeting: 69-84
BRANDT, S. 2005 Erstnachweis der Fisch-Gattung Menaspis EWALD aus dem Kupferschiefer (Zechstein, Ober-Perm) von Gera. Veröffentlichungen des Museum für Naturkunde der Stadt Gera, 32: 157-159, Gera
BRITO, P.M.M. & DEYNAT, P.P. 2004 Freshwater Stingrays from the Miocene of South America with comments on the rise of potamotrygonids (Batoidea, Myliobatiformes). In: ARRATIA, G., WILSON, M. V. H. and CLOUTIER, R. (eds.), Recent advances in the origin and early radiation of vertebrates. Honoring Hans-Peter Schultze: 571-578
BURROW, C.J. & HOVESTADT, D.C. & HOVESTADT-EULER, M. & TURNER, S. & YOUNG, G.C. 2008 Mid-Palaeozoic (Devonian, ?late Emsian) shark McMurdodus whitei from western Queensland, Australia. In: Ginter, M. ed. Warsaw Chondrichthyans symposium, July 29-Aug. 2, abstract. Ichthyolith Issues SP 11, p. 2.
BURROW, C.J. & TURNER, S. 2012 Fish stew: fossil fish taphonomy and the contribution of microfossils in documenting Devonian vertebrate history. In: J.A. Talent ed. Global biodiversity, extinction intervals and biogeographic perturbations through time UNESCO/International Year of Planet Earth ‘life theme'.
CIONE, A.L. 2006 The fishes. In: Fuscaldo, P. and S. Lupo (Eds.). Tell el-Ghaba, a Saite Settlement in North Sinai, Egypt, (Argentine Archaeological Mission 1995-2004). Vol II. Studies. Buenos Aires-CONICET. Formato CD. ISSN 1850 3462, ISBN 978- 950-692-076-0.
DAWSON, T. & DAWSON, L. 2006 Evolution of arid Australia and consequences for vertebrates. In: Merrick, J., Archer, M., Hickey, G., Lee, M. (Eds.), Evolution and Biogeography of Australasian Vertebrates. Auscipub, Sydney, pp. 54-67.
DIDIER, D.A. 2004 Phylogeny and classification of extant Holocephali. In: Carrier, J.C. & Musick, J.A. & Heithaus, M.R. (eds) Biology of Sharks and their Relatives: 115-136
FRIEDMAN, V. 2012 Vertebrate coprolites from the Lower Eagle Ford Group of North Central Texas and their paleoecological significance. New Mexico Museum of Natural History and Science, Bulletin, 57: 221-228
GALLARDO, C. & SHIMADA, K. & SCHUMACHER, B.A. 2011 Fossil marine vertebrates from the lowermost Greenhorn Limestone (Upper Cretaceous: Middle Cenomanian) in southeastern Colorado. Abstract. In: Society for Advancement of Chicanos and Native Americans in Science National Conference, San Jose, California, October 27-30, 2011
GINTER, M. 2005 Ontogenetic development of teeth in Symmoriiform sharks. In: Ivanov, A. & Young, G. (Eds.), Middle Palaeozoic Vertebrates of Laurussia: relationships with Siberia, Kazakhstan, Asia and Gondwana, St. Petersbourg, Russia, August 22-25, 2005, IGCP 491. Ichthyoliths Issues Special Publication, 9: 12-13
GRAF, W. & KLAM, F. 2006 Le systeme vestibulaire : anatomie fonctionnelle et comparee, evolution et developpement. Comptes Rendus - Palevol, 5 (3): 637-655
HAMPE, O. & ABOUSSALAM, Z.S. & BECKER, R.T. 2004 Omalodus teeth (Elasmobranchii: Omalodontida) from the northern Gondwana margin (middle Givetian: ansatus conodont Zone, Morocco). In: ARRATIA, G., WILSON, M. V. H. and CLOUTIER, R. (eds.), Recent advances in the origin and early radiation of vertebrates. Honoring Hans-Peter Schultze: 487-504, 7 fig., 1 tab., 1 App.
IFRIM, C. & FREY, E. & STINNESBECK, W. & BUCHY, M.-C. & GONZÁLEZ GONZÁLEZ, A.H. & LÓPEZ OLIVA, J.G. 2005 Fish assemblage in Lower Turonian carbonates at Vallecillo, N.L., México. Paleos Antiguo, 1: 43-51
IVANOV, A.O. & LUKSEVICS, E. & STINKULIS, G. & TOVMASJAN, K. & ZUPINS, I. & ZABELE, A. 2005 Devonian stratigraphy and vertebrate fauna of the Andoma Hill section (Onega Lake, Russia). Ichthyolith Issues, Special Publication, 9: 17-21.
LUND, R. & GROGAN, E.D. 2004 Five new euchondrocephalan Chondrichthyes from the Bear Gulch Limestone (Serpukhovian, Namurian E2b) of Montana, USA. In: ARRATIA, G., WILSON, M. V. H. and CLOUTIER, R. (eds.), Recent advances in the origin and early radiation of vertebrates. Honoring Hans-Peter Schultze: 505-531
MANGANELLI, G. & BENOCCI, A. & SPADINI, V. 2006 The scientific bibliography of Roberto Lawely (1818-1881) and his contribution to the study of fossil sharks. Archives of Natural History, 33 (2): 267-281
MAO, Y. & MA Q. & FENG Q. 2013 Discovery of Fish Microremains in the Gufeng Formation at the Luojiaba Section from Jianshi, West Hubei. Acta Micropalaeontologica Sinica, 30 (2): 175-183
MARQUET, R. & HERMAN, J. 2009 The Stratigraphy of the Pliocene of Belgium.Palaeofocus, 2: 62p. (incl.7pls.)
RICHTER, M. 2007 First record of Eugeneodontiformes (Chondrichthyes: Elasmobranchii) from the Paraná Basin, Late Permian of Brazil. In: de Souza Carvalho, I., de Cassia Tardin Cassab, R., Schwanke, C., de Araujo Carvalho, M, Sequeira Fernandes, A. C., da Conceição Rodrigues, M. A., Sardenberg Salgado de Carvalho, M., Arai, M. and Queiroz Oliveira, M. E. (eds.), Paleontologia: cenários de vida, 1, 149-156. (Interciéncía Ltda.) Rio de Janeiro.
ROBB, A.J. 2004 Vertebrate fossils from the Upper Cretaceous (Merchantville Formation: Early Campanian) Graham Brickyard locality of New Jersey. Mosasaur, 7: 75-88
SCHINDLER, T. 2011 Das Fossil des Jahres 2011 stammt aus der Pfalz. Haiplatte aus Niederkirchen gewürdigt. Pollichia-Kurier, 27 (3): 30-31, 2 Abb.
STROUGO, A. & CAPPETTA, H. & ELNAHAS, S. 2007 A remarkable Eocene ichthyofauna from the ElGedida glauconitic sandstone, Bahariya oasis, Egypt, and its stratigraphic implications. M.E.R.C. Aïn Shams University, Earth Science Series, 21: 81-98, 4 fig., pl. 1-3.
TURNER, S. 2004 Early vertebrates: analysis from microfossil evidence. In: ARRATIA, G., WILSON, M. V. H. and CLOUTIER, R. (eds.), Recent advances in the origin and early radiation of vertebrates. Honoring Hans-Peter Schultze: 67-94
TURNER, S. & MILLER, R.F. 2008 Protodus jexi, Woodward, 1892 (Chondrichthyes), from the Lower Devonian Campbellton Formation, New Brunswick, Canada. In: Ginter, M. ed. Warsaw Chondrichthyans, abstract. Ichthyolith Issues SP 11, p. 20-21.
YAMAGISHI, H. 2004 Elasmobranch remains from the Taho Limestone (Lower-Middle Triassic) of Ehime Prefacture, Southwest Japan; Mesozoic Fishes 3 - Systematics, Paleoenvironments and Biodiversity: 565-574
Please support www.shark-references.com and send missing papers (not listed papers or papers without the infosymbol) to juergen.pollerspoeck@shark-references.com.
|
|
Meetings:

The KwaZulu-Natal Sharks Board is pleased to confirm that the second Sharks International symposium will be held in Durban, South Africa on 2-6 June 2014. This is a sequel to the highly successful inaugural Sharks International meeting in Cairns, Australia in June 2010. The meeting will comprise four full days of presentations, commencing Monday 2 June and ending Friday 6 June, with Wednesday 4 June set aside for a variety of exciting mid-conference excursions.
The aim of this conference is to provide a forum for the world’s leading shark and ray researchers, along with students and early career scientists to meet, share ideas, update information and report on the progress of their most recent scientific studies. We would encourage any researchers and students with a general interest in the marine environment to also attend as networking opportunities at a conference of this nature and size are enormous.
For further information please visit the Symposium’s web page at:
http://www.sharksinternational.org
Any queries regarding the symposium can also be sent to the organising committee at:
sharksinternational2014@gmail.com |
|

FIRST ANNOUNCEMENT
EEA 2014 European Elasmobranch Association – Annual Scientific Conference 18 years on – Prepared for the Future
The Dutch Elasmobranch Society is proud to host the 18th Annual Scientific Conference of the European Elasmobranch Association from 7th to 9th November 2014 at the Van Hall Larenstein University of Applied Sciences in Leeuwarden, The Netherlands.
Elasmobranchs are increasingly included in high level policy agreements. The conference will provide a platform for those involved in international science and policy and aims to help coordinate the information necessary for the development and implementation of management measures for rays and sharks in European waters. EEA 2014 will be of interest to all those who are involved in the study, management and conservation of chondrichthyans (sharks, skates, rays and chimaeras).
The three-day conference will include theme sessions on policy making, restoration measures, integrated management, husbandry and captive management, tagging and other subjects. The collection, availability and sharing of data will be an underlying theme. There will be plenary talks, a poster session and opportunities for networking and socialising. In addition an excursion will be offered on the last day.
Leeuwarden is a vibrant town with a charming centre and a wide selection of hotels and restaurants. It is the capital city of the northern Province of Fryslân and will be the Cultural Capital of Europe in 2018. There is a twice hourly train service from Schiphol (one direct, one with one connection) and it is easy to reach by road.
See www.elasmobranch.nl/EEA2014 for preliminary details on the conference.
Contact: eea2014@elasmobranch.nl
|
|
New described species/Taxonomic News:
Recent:
STRAUBE, N. & LESLIE, R.W. & CLERKIN, P.J. & EBERT, D.A. & ROCHEL, E. & CORRIGAN, S. & LI, C. & NAYLOR, G.J.P. (2014): On the occurrence of the southern Lanternshark, Etmopterus granulosus, off south Africa, with comments on the validity of E. compagnoi. Deep Sea Research Part II: Topical Studies in Oceanography, in press
Abstract: The Southern Lanternshark, Etmopterus granulosus, is a large species of Lanternshark that has been a source of long-standing taxonomic confusion. Recent work suggests E. granulosus to be conspecific with the New Zealand Giant Lanternshark, Etmopterus baxteri, suggesting that the species may be widespread throughout the Southern Hemisphere. The taxonomic affinity of populations off South Africa, however, has remained uncertain. Herein we show that South African samples are also conspecific with E. granulosus based on both molecular and morphometric data. These results extend the known distribution range of this species to South Africa and the southern Indian Ocean, strengthening the hypothesis that E. granulosus has a circum-Antarctic distribution. In addition we show that there is a cryptic, granulosus-like species in South African waters that can likely be assigned to Etmopterus compagnoi.
Fossil:
COOK, T.D. & EATON, J.G. & NEWBREY, M.G. & WILSON, M.V.H. (2014): A new genus and species of freshwater stingray (Myliobatiformes, Dasyatoidea) from the latest middle Eocene of Utah, U.S.A. Journal of Paleontology, 88 (3): 497-503
New genus: Saltirius
New species: Saltirius utahensis
Abstract: Teeth of a new freshwater dasyatoid ray recovered from the latest middle Eocene Brian Head Formation of southern Utah represent the youngest freshwater stingray so far known in the fossil record of North America. The crown morphology of Saltirius utahensis n. gen. n. sp. exhibits strong sexual dimorphism, with the presumed males bearing two prominent margino-labial protuberances and a bifid cusp that produces a saltire-like outline. This unique crown separates this genus and species from any known extinct or extant myliobatiform, but does have some resemblance to the crown of Asterotrygon maloneyi from the lower Eocene Green River Formation of Wyoming. The occurrence of S. utahensis in the Brian Head Formation provides additional evidence for the persistence of warm subtropical temperatures during the late Eocene in southern Utah.
PAULIV, V.E. & DIAS, E.V. & SEDOR, F.A. & RIBEIRO, A.M. (2014): A new Xenacanthiformes shark (Chondrichthyes, Elasmobranchii) from the late Paleozoic Rio do Rasto formation (paraná basin), southern Brazil. Anais da Academia Brasileira de Ciencias, 86 (1): 135-145
New species: Xenacanthus ragonhai
Abstract: The Brazilian records on Xenacanthiformes include teeth and cephalic spines from the Parnaíba, Amazonas and Paraná basins. This work describes a new species of Xenacanthidae, collected in an outcrop of Serrinha Member of Rio do Rasto Formation (Wordian to Wuchiapingian), Paraná Basin, municipality of Jacarezinho, State of Paraná. The teeth of the new species are two or three-cuspidated and the aboral surface show a smooth concavity and one rounded basal tubercle. The coronal surface presents one semi-spherical and subcircular coronal button, and also two lateral main cusps and one central (when present) with less than one fifth of the size of the lateral cusps in the labial portion. The lateral cusps are asymmetric or symmetric, rounded in transversal section, lanceolate in longitudinal section, devoid of lateral carinae and lateral serrations, and with few smooth cristae of enameloid. In optical microscope the teeth show a trabecular dentine (osteodentine) base, while the cusps are composed by orthodentine, and the pulp cavities are non-obliterated by trabecular dentine. The fossil assemblage in the same stratigraphical level and in the whole Rio do Rasto Formation indicates another freshwater record for xenacanthid sharks.
WELTON, B.J. (2014): A new fossil basking shark (Lamniformes: Cetorhinidae) from the Middle Miocene Sharktooth Hill Bonebed, Kern County, California. Contributions in Science, 522: 29-44
New species: Cetorhinus huddlestoni
Abstract: Isolated teeth of a middle Miocene cetorhinid genus, Cetorhinus Blainville 1816, occur abundantly in rocks of the Round Mountain Silt, Sharktooth Hill Bonebed, southeastern San Joaquin Valley, Kern County, California. Tooth sets of juvenile and adult dentitions of the Sharktooth Hill Cetorhinus were reconstructed and used as a basis for description of a new species, C. huddlestoni. The teeth of C. huddlestoni are of about the same size as those of adult C. maximus, the only other species of Cetorhinus. Cetorhinus huddlestoni has weak gradient monognathic heterodonty, moderate dignathic heterodonty, no disjunct monognathic heterodonty in either jaw, and strong ontogenetic heterodonty; there is presently no evidence of sexual dimorphism. Attributes noted in C. huddlestoni and absent in C. maximus include the following: bilobate roots in most adult teeth; broad based crowns with a wide, tall cusp and sharp apex in adult teeth; narrow and erect crowns in juvenile teeth; a smooth mesial and distal profile across the crown foot–root junction; mesial cusplets on some teeth from distal tooth rows in juveniles, and the sporadic occurrence of a bladelike mesial cusplet in some adult teeth; long mesial cutting ridges in adult teeth. Relative to C. huddlestoni, the teeth of C. maximus appear to be more reduced, in having narrower cusps with rounded or blunt apices in adults; much higher lingual necks; hooklike, nearly horizontal cusps in juveniles; and significantly more robust, bulbous roots, perhaps to compensate for the loss of root lobes. Cetorhinus huddlestoni is presently only known from the eastern North Pacific, middle Miocene Round Mountain Silt, Sharktooth Hill Bonebed, and most closely resembles Cetorhinus cf. C. maximus from the early Pliocene Sands of Kattenjijk near Kallo, Belgium.
|
|
New Paper
Recent Papers:
ALAMA, M.T. & PETIT, R.A. & READA, T.D. & DOVE, A.D.M. (2014): The complete mitochondrial genome sequence of the world's largest fish, the whale shark (Rhincodon typus), and its comparison with those of related shark species. Gene, 539 (1): 44-49 http://dx.doi.org/10.1016/j.gene.2014.01.064
AL-SHAJIBI, S.R. & CHESALIN, M.V. & AL-SHAGAA, G.A. (2014): New Records of Sharks from Southern Coastal Waters of Oman in the Arabian Sea. Pakistan Journal of Zoology, 46 (1): 281-284
BECKMANN, C.L. & MITCHELL, J.G. & STONE, D.A.J. & HUVENEERS, C. (2014):Inter-Tissue Differences in Fatty Acid Incorporation as a Result of Dietary Oil Manipulation in Port Jackson Sharks (Heterodontus portusjacksoni). Lipids, in presshttp://dx.doi.org/10.1007/s11745-014-3887-6
CHEN, X. & PENG, X. & ZHANG, P. & YANG, S. & LIU, M. (2014): Complete mitochondrial genome of the spadenose shark (Scoliodon macrorhynchos).Mitochondrial DNA, 25 (2): 91-92 http://dx.doi.org/10.3109/19401736.2013.784751
CHEN, X. & ZHOU, Z. & PICHAI, S. & HUANG, X. & ZHANG, H. (2014): Complete mitochondrial genome of the brownbanded bamboo shark Chiloscyllium punctatum.Mitochondrial DNA, 25 (2): 113-114 http://dx.doi.org/10.3109/19401736.2013.786710
CLARKE, T.M. & ESPINOZA, M. & WEHRTMANN, I.S. (2014): Reproductive ecology of demersal elasmobranchs from adata-deficient fishery, Pacific of Costa Rica, Central America. Fisheries Research, 157: 96-105 http://dx.doi.org/10.1016/j.fishres.2014.04.003
CLUA, E. & BESCOND, P.-M. & REID, D. (2014): Fatal attack by a juvenile tiger shark, Galeocerdo cuvier, on a kitesurfer in New Caledonia (South Pacific). Journal of Forensic and Legal Medicine, in press http://dx.doi.org/10.1016/j.jflm.2014.04.005
DIAS, J.F. & FERNANDEZ, W.S. & SCHMIDT, T.C.S. (2014): Length-weight relationship of 73 fish species caught in the southeastern inner continental shelf region of Brazil. Latin American Journal of Aquatic Research, 42 (1): 127-136http://dx.doi.org/103856/vol42-issue1-fulltext-10
EBERT, D.A. & KNUCKEY, J.D.S. & KAMIKAWA, D.J. (2014): First eastern North Pacific record of the velvet dogfish, Zameus squamulosus (Chondrichthyes: Squaliformes: Somniosidae). Marine Biodiversity Records, 7: e48http://dx.doi.org/10.1017/S1755267214000499
FARIA, V.V. & CHIARAMONTE, G.E. & OIJEN, M.J.P. (2014): Publication date and author spelling for the hidden angelshark Squatina occulta. Journal of Fish Biology, 84 (4): 1185-1187 http://dx.doi.org/10.1111/jfb.12325
FLAMMANG, B.E. (2014): The fish tail as a derivation from axial musculoskeletal anatomy: an integrative analysis of functional morphology. Zoology, 117 (1): 86-92http://dx.doi.org/10.1016/j.zool.2013.10.001
FRANKLIN, O. & PALMER, C. & DYKE, G. (2014): Pectoral fin morphology of batoid fishes (Chondrichthyes: Batoidea): Explaining phylogenetic variation with geometric morphometrics. Journal of Morphology, in press http://dx.doi.org/10.1002/jmor.20294
GERAGHTY, P.T. & WILLIAMSON, J.E. & MACBETH, W.G. & BLOWER, D.C. & MORGAN, J.A.T. & JOHNSON, G. & OVENDEN, J.R. & GILLINGS, M.R. (2014):Genetic structure and diversity of two highly vulnerable carcharhinids in Australian waters. Endangered Species Research, 24 (1): 45-60 http://dx.doi.org/10.3354/esr00580
GOWTHAMAN, A.M. & JAWAHAR, P. & VENKATARAMANI, V.K. (2014): New occurrence of big eye thresher shark Alopias superciliosus Lowe, 1841 in Gulf of Mannar, southeast coast of India. Indian Journal of Geo-Marine Sciences, 43 (5): 883-885
HIGGS, N.D. & GATES, & A.R. & JONES, D.O.B. (2014): Fish Food in the Deep Sea: Revisiting the Role of Large Food-Falls. PLoS ONE, 9 (5): e96016http://dx.doi.org/10.1371/journal.pone.0096016
HORVAT, M. & DEGENEK, N. & LIPEJ, L. & TRATNIK, J.S. & FAGANELI, J. (2014): Trophic transfer and accumulation of mercury in ray species in coastal waters affected by historic mercury mining (Gulf of Trieste, northern Adriatic Sea).Environmental Science and Pollution Research, 21 (6): 4163-4176http://dx.doi.org/10.1007/s11356-013-2262-0
KADRI, H. & MAROUANI, S. & BRADAÏ, M.N. & BOUAÏN, A. & MORIZE, E. (2014):Distribution and morphometric characters of the Mediterranean thornback ray, Raja clavata (Chondrichthyes: Rajidae) in the Gulf of Gabès (Tunisia, Central Mediterranean). [Distribution et caractères morphométriques de la raie bouclée de la Méditerranée, Raja clavata (Chondrichthyens : Rajidae) dans le golfe de Gabès (Tunisie, Méditerranée centrale)]. Marine Life, 18: 9-16
KIZHAKUDAN, S.J. & RAJAPACKIAM, S. (2013): First report of the crocodile shark Pseudocarcharias kamoharai (Matsubara,1936)from Chennai, southeast coast of India.Journal of the Marine Biological Association of India, 55 (1): http://dx.doi.org/10.6024/jmbai.2013.55.1.01734-14
LE BOURG, B. & KISZKA, J. & BUSTAMANTE, P. (2014): Mother–embryo isotope (δ15N, δ13C) fractionation and mercury (Hg) transfer in aplacental deep-sea sharks.Journal of Fish Biology, 84 (5): 1574-1581 http://dx.doi.org/10.1111/jfb.12357
MARCENIUK, A.P. & CAIRES, R. & WOSIACKI, W.B. & DI DARIO, F. (2014):Conhecimento e conservação dos peixes marinhos e estuarinos (Chondrichthyes e Teleostei) da costa norte do Brasil. [Knowledge and conservation of the marine and estuarine fishes (Chondrichthyes and Teleostei), of the north coast of Brazil.] Biota Neotropica, 13 (4): 251-259
MEJÍA-FALLA, P.A. & CORTÉS, E. & NAVIA, A.F. & ZAPATA, F.A. (2014): Age and Growth of the Round Stingray Urotrygon rogersi, a Particularly Fast-Growing and Short-Lived Elasmobranch. PLoS ONE, 9 (4): e96077 http://dx.doi.org/10.1371/journal.pone.0096077
NAVARRO, J. & LÓPEZ, L. & COLL, M. & BARRÍA, C. & SÁEZ-LIANTE, R. (2014):Short- and long-term importance of small sharks in the diet of the rare deep-sea shark Dalatias licha. Marine Biology, in press http://dx.doi.org/10.1007/s00227-014-2454-2
PAIG-TRAN, E.W.M. & SUMMERS, A.P. (2014): Comparison of the Structure and Composition of the Branchial Filters in Suspension Feeding Elasmobranchs.Anatomical Record, 297 (4): 701-715 http://dx.doi.org/10.1002/ar.22850
PÉREZ-JIMÉNEZ, J.C. (2014): Historical records reveal potential extirpation of four hammerhead sharks (Sphyrna spp.) in Mexican Pacific waters. Reviews in Fish Biology and Fisheries, in press http://dx.doi.org/10.1007/s11160-014-9353-y
RAMACHANDRAN, S. & AYOOB, A.E. & KOYA, P.P. (2014): A new distributional record of bramble shark Echinorhinus brucus (Bonnaterre, 1788) from the seamount of Lakshadweep Islands with description on skeletal rings. Journal of the Marine Biological Association of India, 55 (2): 5-11 http://dx.doi.org/10.6024/jmbai.2013.55.2.01787-0x
RANDHAWA, H. & POULIN, R. & KRKOSEK, M. (2014): Increasing rate of species discovery in sharks coincides with sharp population declines: implications for biodiversity. Ecography, in press http://dx.doi.org/10.1111/ecog.00793
RITTER, E.K. & COMPAGNO, L.J.V. (2013): First record of a smalltooth sandtiger shark, Odontaspis ferox, from the Galapagos Islands. Marine Biodiversity Records, 6: e130 http://dx.doi.org/10.1017/S1755267213001115
SCHILLER, L. & ALAVA, J.J. & GROVE, J. & RECK, G. & PAULY, D. (2014): The demise of Darwin's fishes: evidence of fishing down and illegal shark finning in the Galápagos Islands. Aquatic Conservation: Marine and Freshwater Ecosystems, in press http://dx.doi.org/10.1002/aqc.2458
SIMPSON, M.R. & MIRI, C.M (2014): A pre-COSEWIC assessment of Porbeagle Shark (Lamna nasus) in Newfoundland and Labrador waters. DFO Canadian Science Advisory Secretariat, Research Document 2013/088: 19 p.
STRAUBE, N. & LESLIE, R.W. & CLERKIN, P.J. & EBERT, D.A. & ROCHEL, E. & CORRIGAN, S. & LI, C. & NAYLOR, G.J.P. (2014): On the occurrence of the southern Lanternshark, Etmopterus granulosus, off south Africa, with comments on the validity of E. compagnoi. Deep Sea Research Part II: Topical Studies in Oceanography, in press http://dx.doi.org/10.1016/j.dsr2.2014.04.004
THIEL, R. & WINKLER, H. & BÖTTCHER, U. & DÄNHARDT, A. & FRICKE, R. & GEORGE, M. & KLOPPMANN, M. & SCHAARSCHMIDT, T. & UBL, C. & VORBERG, R. (2013): Rote Liste und Gesamtartenliste der etablierten Fische und Neunaugen (Elasmobranchii, Actinopterygii & Petromyzontida) der marinen Gewässer Deutschlands. Naturschutz und Biologische Vielfalt, 70 (2): 11-76, Bundesamt für Naturschutz, Bonn
VIANNA, G.M.S. & MEEKAN, M.G. & BORNOVSKI, T.H. & MEEUWIG, J.J. (2014):Acoustic Telemetry Validates a Citizen Science Approach for Monitoring Sharks on Coral Reefs. PLoS ONE, 9 (4): e95565 http://dx.doi.org/10.1371/journal.pone.0095565
VIGNAUD, T.M. & MAYNARD, J.A. & LEBLOIS, R. & MEEKAN, M.G. & VÁZQUEZ-JUÁREZ, R. & RAMÍREZ-MACÍAS, D. & PIERCE, S.J. & ROWAT, D. & BERUMEN, M.L. & BEERAVOLU, C. & BAKSAY, S. & PLANES, S. (2014): Genetic structure of populations of whale sharks among ocean basins and evidence for their historic rise and recent decline Molecular Ecology, 23 (10): 2590–2601http://dx.doi.org/10.1111/mec.12754
WATERS, J.D. & COELHO, R. & FERNANDEZ-CARVALHO, J. & TIMMERS, A.A. & WILEY, T. & SEITZ, J.C. & MCDAVITT, M.T. & BURGESS, G.H. & POULAKIS, G.R. (2014): Use of encounter data to model spatio-temporal distribution patterns of endangered smalltooth sawfish, Pristis pectinata, in the western Atlantic. Aquatic Conservation: Marine and Freshwater Ecosystems, in presshttp://dx.doi.org/10.1002/aqc.2461
WEN, L. & WEAVER, J.C. & LAUDER, G.V. (2014): Biomimetic shark skin: design, fabrication and hydrodynamic function. Journal of Experimental Biology, 217: 1656-1666 http://dx.doi.org/10.1242/jeb.097097
Parasitology:
AUSTIN, C.M. & TAN, M.H. & LEE, Y.P. & CROFT, L.J. & MEEKAN, M.G. & PIERCE, S.J. & GAN, H.M. (2014): The complete mitogenome of the whale shark parasitic copepod Pandarus rhincodonicus Norman, Newbound & Knott (Crustacea; Siphonostomatoida; Pandaridae) - a new gene order for the copepoda. Mitochondrial DANN, in press http://dx.doi.org/10.3109/19401736.2014.913147
Fossil:
COOK, T.D. & EATON, J.G. & NEWBREY, M.G. & WILSON, M.V.H. (2014): A new genus and species of freshwater stingray (Myliobatiformes, Dasyatoidea) from the latest middle Eocene of Utah, U.S.A. Journal of Paleontology, 88 (3): 497-503http://dx.doi.org/10.1666/13-046
DIEDRICH, C.G. (2014): Skeleton of the Fossil Shark Isurus denticulatus from the Turonian (Late Cretaceous) of Germany—Ecological Coevolution with Prey of Mackerel Sharks. Paleontology Journal, 2014: ID 934235 http://dx.doi.org/10.1155/2014/934235
GORMAN, K.L. & SHIMADA, K. & WITZKE, B.J. (2014): Late Cretaceous Marine Fishes from the Basal Greenhorn Limestone in Western Iowa. Transactions of the Kansas Academy of Science, 117 (1-2): 91-99 http://dx.doi.org/10.1660/062.117.0114
OTERO, R.A. & GUTSTEIN, C.S. & VARGAS, A. & RUBILAR-ROGERS, D. & YURY-YAÑEZ, R. & BASTÍAS, J. & RAMÍREZ, C. (2014): New chondrichthyans from the Upper Cretaceous (Campanian–Maastrichtian) of Seymour and James Ross islands, Antarctica. Journal of Paleontology, 88 (3): 411-420http://dx.doi.org/10.1666/13-041
PAULIV, V.E. & DIAS, E.V. & SEDOR, F.A. & RIBEIRO, A.M. (2014): A new Xenacanthiformes shark (Chondrichthyes, Elasmobranchii) from the late Paleozoic Rio do Rasto formation (paraná basin), southern Brazil. Anais da Academia Brasileira de Ciencias, 86 (1): 135-145 http://dx.doi.org/10.1590/0001-37652014107612
TRIKOLIDI, F.A. (2014): Cow sharks (Hexanchiformes) from the Cretaceous deposits of the Crimea. (In russian) Proceedings of the Zoological Institute, Russian Academy of Sciences, 318 (1): 76-97
WELTON, B.J. (2014): A new fossil basking shark (Lamniformes: Cetorhinidae) from the Middle Miocene Sharktooth Hill Bonebed, Kern County, California. Contributions in Science, 522: 29-44
WRIGHT, J.K. & BASSETT-BUTT, L. & COLLINSON, M. (2014): Fatally bitten ammonites from the Lower Calcareous Grit Formation (Upper Jurassic) of NE Yorkshire, UK. Proceedings of the Yorkshire Geological Society, in presshttp://dx.doi.org/10.1144/pygs2014-336
|
|
The Manzano Ctenacanth: Godzilla Shark of New Mexico!
The Story

About 300 million years ago, in the Manzano Mountains east of Albuquerque, a shallow warm lagoonal estuary was home to a wide variety of plants and animals. Environmental conditions of the bottom of this lagoonal estuary suggest oxygen was extremely poor or not present. This prevented bacterial decomposition of organisms that died and sank to the fine- grained silts and muds to be preserved in relative completeness and in high detail. These conditions preserved thin plant cells, bone, cartilage, and soft tissues.
Paleontologists and geologists from the Smithsonian, Carnegie Museum, University of Kansas and the New Mexico Museum of Natural History and Science have researched the exquisitely preserved fossils from the Manzano Mountains since the late 1960’s. Fossils of plants, invertebrates, small fish, and small amphibians have been collected over the years.
In 2013, during a paleontological conference day trip to the Manzano Mountains, shark paleontologist John-Paul Hodnett serendipitously came across the tip of the nose of a nine foot long ctenacanth (TEEN-uh-kanth) shark (Figure 1). Ctenacanth sharks have large spines in front of their dorsal fins and first appeared in the fossil record during the Late Devonian, about 380 million years ago, and survived until the Early Cretaceous, about 135 million years ago. Scientists in the past proposed that ctenacanths were direct ancestors of modern sharks. Current research, however, suggests ctenacanths might be more closely related to the extinct basal sharks rather than to their modern kin. Until this recent discovery in New Mexico, ctenacanth sharks were known only from two partial skeletons, a few partial skulls, and isolated teeth and dorsal spines. For a shark paleontologist the 2013 discovery was a dream come true…
The shark was exposed on its right side in a fine grained limestone. Due to its size the shark was removed in three sections to be preserved and re-assembled in the paleontology laboratory at the New Mexico Museum of Natural History and Science. The top of the braincase and the right and left upper jaws of the skull, the dorsal fin spines, the vertebral column, and the pectoral and pelvic fins were exposed with mechanical preparation by museum staff. The tail is still being prepared. It is currently under study by Hodnett, Dr. Spencer G. Lucas, and scientists from the American Museum of Natural History in New York and Northern Arizona University in Flagstaff.

What We Are Doing
- Using Computed Tomography (CT), we are scanning the head section of the “Manzano Ctenacanth”
- Scanning the head section we are looking at the composition of the braincase, the palatoquadrate (a single cartilage structure that is the cheek and upper jaw), and looking for the cartilages that make up the Meckel’s cartilages (the lower jaws) and the gill arches, which are not initially seen on the surface of the rock containing the specimen.
- We are CT scanning the full body skeleton of Cobelodus, a smaller shark and a relative of the “Manzano Ctenacanth” found in the Manzano Mountains (Figure 1)
- For Cobelodus, we are looking for hidden structures that are hard to see or that are hidden by sediments to determine the morphology of the skull, visceral skeleton, the body column, and the fins
What We Know
- The “Manzano Ctenacanth” is presently the largest and most complete ctenacanth in the world
- It is presently the largest vertebrate fossil to be discovered at the fossil site in the Manzano Mountains
- It is approximately 85 to 90+% complete
- The lack of “claspers”, a skeletal sexual organ on the pelvic fins of male sharks, makes the “Manzano Ctenacanth” a female
- The size of the dorsal fin spines (which are almost two feet long) and the shape of the teeth suggest the “Manzano Ctenacanth” is a new species
- The body of the “Manzano Ctenacanth” was covered with small tooth-like scales, called dermal denticles
- Cobelodus is the most common shark from the fossil site in the Manzano Mountains
- Most of the specimens of Cobelodus are small, smaller than “adult specimens” from a similar aged locality in Indiana. These specimens have been proposed to be juveniles in the past.
Why CT Scanning Is Important
- Knowing the structure of the braincase and the visceral skeleton (jaws and gill arches) in the “Manzano Ctenacanth” and Cobelodus will help determine the relationship between these sharks to other extinct and modern cartilaginous fishes (Figure 2)
- The completeness of the “Manzano Ctenacanth” will allow us to use it as a keystone specimen to determine the missing data of other ctenacanth shark specimens around the world
- The “Manzano Ctenacanth”skull can be recreated in three dimensions from the CT data
- A three dimensional skull model can be used to determine how the jaws and gills operated and can help determine how strong of a bite force the “Manzano Ctenacanth” could produce
- A 3D skull of the “Manzano Ctenacanth” could be “printed” for study and display
- CT scanning the Cobelodus will allow us to test the hypothesis that the New Mexico sample represents juveniles in a marine nursery, or are small adults of a new species of Cobelodus, or a completely new shark
- The full skeleton of Cobelodus can be reconstructed three dimensionally from the CT data
- A skeletal model can be made to determine how the shark moved and hunted
- A 3D skeleton of Cobelodus could be “printed” for study and display

Figure 1: bottom left, outline drawing of the “Manzano Ctenacanth”NMMNH P-68537 exposed to the surface; top left, reconstruction of the “Manzano Ctenacanth”; bottom right, outline drawing of NMMNH P-19182, a Cobelodusshark, from New Mexico Quarry; top right, reconstruction of Cobelodus . Both illustrations drawn to scale to a 5’10” male human (©New Mexico Museum of Natural History and Science/JP Hodnett)

Figure 2: “Shark Family Tree” by Ray Troll showing the branch where ctenacanth sharks are related to other cartilaginous fishes and “Bony” fishes (© Ray Troll, reproduced with permission, http://www.trollart.com/)
© Spencer G. Lucas, Ph.D., New Mexico Museum of Natural History and Science, 1801 Mountain Road N. W., Albuquerque, New Mexico 87104-1375 USA
|
|
source: http://theconversation.com/scientists-focusing-on-the-wrong-sharks-in-the-wrong-places-26245
Scientists focusing on the wrong sharks in the wrong places
 Blacktip whaler, a heavily targeted species for shark fisheries. Rob Harcourt Blacktip whaler, a heavily targeted species for shark fisheries. Rob Harcourt
Sharks are in danger in most parts of the world, with a quarter of all sharks and rays now threatened with extinction. This ongoing collapse of shark populations has already hadfar-reaching effects on marine ecosystems. To arrest this trend we desperately need scientific data that effectively inform conservation management. But are we focusing our research effort in the right areas?
We looked at 20 years' worth of scientific papers on shark conservation to see if shark science is meeting the needs of conservation. Our results, included in a forthcoming book, suggest that the species most in need, and the places where most damage is done, aren’t getting the attention they deserve.
Are we looking in the right places?
For science to have the best chance of helping to protect sharks, it should be focused on places where the need for conservation is greatest. As fishing is the most serious threat to sharks, research efforts should therefore correspond to the places with the most fishing.
Pleasingly, our study found that although shark fishing has increased, scientific effort has also increased substantially in the past few decades. In 1992 only 2 peer-reviewed studies specifically on shark conservation and management were published, by 2011, 80 were published. However, the increase in scientific output is largely driven by two countries: Australia and the United States. In the countries where most shark landings occur (Indonesia, India, Taiwan and Spain) there is almost no research on shark conservation or management.
 National contributions to shark landings (above) and scientific output (below) (source: shark landing data were obtained from the FAO).Modified from Figure 5.2 in : Momigliano, P. and Harcourt, Robert (in press). The Science-Law Disconnect. In: Klein, N. and Techera, E. (eds.) Sharks: Conservation, Governance and Management. Earthscan Series, Routledge National contributions to shark landings (above) and scientific output (below) (source: shark landing data were obtained from the FAO).Modified from Figure 5.2 in : Momigliano, P. and Harcourt, Robert (in press). The Science-Law Disconnect. In: Klein, N. and Techera, E. (eds.) Sharks: Conservation, Governance and Management. Earthscan Series, Routledge
This lack of research effort is mirrored by the failure of these countries to implement national shark management plans along United Nations guidelines. There is therefore very little information on the conservation status of some of the world’s most vulnerable sharks, and few plans in place to improve the prospects of these populations.
Can we change the situation?
This geographic bias can be at least partly explained by wealth. Research is expensive, and countries that catch the most sharks are relatively poor. But as sharks are key players in marine ecosystems all over the world, we all have a vested interest in their conservation.
Moreover, many shark species are highly mobile. Sharks caught in one country’s waters may well have spent much of their lives elsewhere. That means that effective conservation in one region can be spoiled by poor management in a neighbouring country.
We suggest that countries that already invest heavily in conservation and management research have a powerful incentive to cooperate with those that cannot afford it.
For example, Indonesia catches more sharks than any other country in the world. If Australia were to focus some of its shark research on its nearest neighbour, the collaboration could deliver big gains in sustainable fishing management.
 Carcasses of requiem sharks at a landing site in Indonesia. Reporting official landings of approximately 100 000 tons of sharks per year, Indonesia is the leading shark fishing nation in the world. Copyright: Vanessa Jaiteh Carcasses of requiem sharks at a landing site in Indonesia. Reporting official landings of approximately 100 000 tons of sharks per year, Indonesia is the leading shark fishing nation in the world. Copyright: Vanessa Jaiteh
Are we doing the most effective science?
Australia publishes more studies on shark conservation research, per capita, than any other country. But we cannot afford to be complacent about the type of science that we do. Our study found major biases in which species of shark attracted the attention of researchers.
If conservation research is to be useful, it should prioritise species that are most at risk. Yet of 479 scientific articles on shark conservation and/or management published worldwide over the past 20 years, only 22 (less than 5%) focused on globally endangered species. And 16 of those were focused on a single species that is also of economic importance: the scalloped hammerhead. That leaves just six papers (on four species) in two decades which looked at all of the other globally endangered sharks (24 species).
More than 200 species (over 40%) of sharks are classified by the International Union for the Conservation of Nature as “data deficient”: we simply do not have enough data to make a call as to whether they are endangered or not. Yet, in the past two decades only 30 studies focusing on 14 data deficient species have been published. Many of these species are likely to be threatened or endangered but without hard data we just don’t know, so effective management is impossible.
Specifically referring to shark conservation, of the 20 most studied species, only the scalloped hammerhead is endangered and just two are listed as data deficient. We found that the vast majority of research is highly skewed towards species that are either economically important (such as the blacktip shark), or charismatic, such as the whale shark and the great white shark.
This skew may be explained by the advantages charismatic species confer in leveraging funding, combined with evidence that publishing in high-impact journals is easier with popular species, with obvious implications for researchers' careers.
Importantly, bycatch species are largely ignored, despite the fact that most sharks are caught as bycatch. This has serious consequences. In southeastern Australia, for example, no data on bycatch of deepwater sharks were available for many decades. By the time this data became available, some species – such as the now critically endangered Harrison’s dogfish – had declined by nearly 99%.
We have a long way to go to improve shark conservation science. While the research effort has increased over the last 20 years, it has done so in only a few countries and for a handful of species – and not always those most in need in either case.
If we want to ensure that sharks are properly managed right around the world, we need to start paying closer attention to the “where” and “what” of shark science.
|
|
source: http://deepseanews.com/2014/05/dead-elasmobranchs-on-the-seafloor-are-not-as-appetizing-as-one-might-assume/
Dead Elasmobranchs on the Seafloor are Not as Appetizing as One Might Assume
 Most fauna in the deep-sea rely upon a drizzle of particles of decaying animals and feces. This marine snow is of low food quality as you might expect death and feces to be. Occasionally, deep-sea buffets occur in the form of a large food fall, a nice way to a near complete carcass. My work on chunks of wood on the deep-sea floor represents one type of these smorgasbords. Other large food falls occur in the form of the well-known whale falls. Natural food falls, i.e. scientist not tossing wood or a whales into the deep ocean, are rarely encountered. Only nine vertebrate carcasses have ever been documented on the seafloor. Add to that four more thanks to Nicholas Higgs, Andrew Gates, and Daniel Jones. Most fauna in the deep-sea rely upon a drizzle of particles of decaying animals and feces. This marine snow is of low food quality as you might expect death and feces to be. Occasionally, deep-sea buffets occur in the form of a large food fall, a nice way to a near complete carcass. My work on chunks of wood on the deep-sea floor represents one type of these smorgasbords. Other large food falls occur in the form of the well-known whale falls. Natural food falls, i.e. scientist not tossing wood or a whales into the deep ocean, are rarely encountered. Only nine vertebrate carcasses have ever been documented on the seafloor. Add to that four more thanks to Nicholas Higgs, Andrew Gates, and Daniel Jones.
Off the Angolan African coast, these researchers document one whale shark and three ray carcasses at 1200 meters on the seafloor. This is the first time any of these have been documented as deep-sea food falls and only recently have living whale sharks even been documented off Angola.
Despite one of the carcasses being covered in 54 eelpouts, a considerable amount of flesh still existed on the carcasses.
[in prior studies] When presented with elasmobranch and tuna bait on a baited camera trap, scavengers clearly preferred tuna and only consumed the elasmobranch once the tuna was gone…Repeated experiments in this region using [bony] fish as bait showed a 10-fold increase in scavenging rates compared to that when elasmobranch was used.
So why in a food desert like the deep sea would fresh meat not be consumed quickly? Apparently, elasmobranch, i.e. shark and ray, flesh is bit unpalatable and tough to chew. The tough, sand-paper-skin may prove a formidable barrier to scavenger jaws. The high ammonia content of elasmobranch flesh may also be, to say the least, unappetizing. The carcasses may also smell like death and deter other scavenging elasmobranchs.
Other uncharacterized chemicals that are found in rotting elasmobranch flesh (necromones) have been proven to strongly deter shark scavenging and invoke an alarm response, even among different species of elasmobranch. If this phenomenon extends to deep-sea scavenging elasmobranchs, it can be assumed that the Portugese dogfish, Centroscymnus coelolepis, would have been deterred from scavenging the elasmobranch carcasses. This will have severely hindered utilization of the carcasses by other species, since C. coelolepis is the dominant scavenger off the Angola margin.
Yet despite the smell of death and urine, a dead elasmobranch still provides an essential snack in the deep sea. The researchers estimate that these elasmobranchs represent 4% of the total amount food that sinks to the seafloor off Angola in the form of marine snow.
Higgs, N., Gates, A., & Jones, D. (2014). Fish Food in the Deep Sea: Revisiting the Role of Large Food-Falls PLoS ONE, 9 (5) DOI: 10.1371/journal.pone.0096016
|
|
source: http://www.cbc.ca/news/canada/nova-scotia/endangered-porbeagle-sharks-still-at-risk-say-scientists-1.2637129
Endangered porbeagle sharks still at risk, say scientists
Recent DFO estimates put the population at about a quarter of what it was in 1960s

The fierce-looking shark is sometimes confused with its larger relative, the great white shark, due to its similar body profile. (Steven Campana/BIO)
A decade after it was declared an endangered species, a group of Canadian scientists says the porbeagle shark remains at risk.
This comes after a re-examination of the species sometimes called “Canada's shark” because it spends most of its life swimming in Canadian waters.
Recent estimates from the Department of Fisheries and Oceans put the population at about a quarter of what it was in 1960s — before overfishing devastated stocks.
The Committee on the Status of Endangered Wildlife in Canada (COSEWIC) re-examined the porbeagle’s endangered status last week in Halifax — and this week re-confirmed it.
“To be reassessed after ten years shows there has been very little progress in the rebuilding of this very vulnerable species,” said Katie Schliet, with the Ecology Action Centre.

Top Canadian shark researcher Steven Campana, pictured here tagging a shark, said there's cause for hope. (Supplied)
Globally, shark populations are on the decline. According to COSEWIC, the porbeagle’s low fertility rates and late maturity make it “extremely vulnerable” to overfishing and capture as bycatch.
Direct fishing for porbeagles was stopped in Canada in 2013. But according to the committee, there’s still a bycatch of unknown magnitude in Canadian waters and there's no way to know how many are being taken in international waters.
However, top Canadian shark researcher Steven Campana, a senior scientist at the Bedford Institute of Oceanography, said there's cause for hope.
“It’s recovering now, slowly. It’s going to take time but it is recovering,” he said.
Campana has spent years studying porbeagle sharks. He said, in recent years, fishermen are catching and mostly releasing more porbeagle as a bycatch.
“They’re catching far larger numbers than they ever did before. This applies — not just in Nova Scotia — but also up in Newfoundland, for instance. So, certainly, everything that I'm getting is suggesting that there is quite a few of them out there,” he said.
Campana estimates the North Atlantic population at 300,000 — most of those are in Canadian waters. Those numbers may seem high, but Campana urges caution.
“You know, that’s a lower number than we would like — let’s make no mistake. Even though I’ve got some optimism for the population, it should still be, probably, twice as high,” he said.
The fierce-looking shark is sometimes confused with its larger relative, the great white shark, due to its similar body profile.
However, swimmers need not worry. Although porbeagles are found in waters surrounding Atlantic Canada, they eat primarily fish and squid.
Environmentalists say more needs to be done, including an outright ban on landing any porbeagles ashore.
DFO said it will continue to monitor the porbeagle but bigger species, like the basking shark, are at even a greater risk.
|
|
source: http://www.independent.co.uk/environment/conservationists-rejoice-as-cornwall-is-awash-with-basking-sharks-9346820.html
Conservationists rejoice as Cornwall is awash with basking sharks
So many basking sharks have already been spotted in British waters that experts are declaring this the best start to shark season in living memory.
A wildlife tour group reported sighting 19 basking sharks up to 25 feet in length last weekend as the eight-tonne travellers begin to arrive off the south west coast.
The animals, which travel to temperate waters and can stay in British regions until October, have been growing in numbers year on year according to The Shark Trust. A total of 266 Basking Shark sightings were reported to the Trust last year as it hopes for an even higher number in 2014.
“To see so many this early has been an absolute honour and it is exciting to consider what the rest of the season may hold for us,” said Captain Keith Leeves, a veteran skipper with AK Wildlife Cruises, told the Western Morning News.
“We have been blown away with the size of the sharks too, with several sharks being over 20 feet long, which is something truly special to behold! This has been one of the best starts to a shark season in living memory.”
Crew Member Billy Burton said: “Guests have been absolutely blown away by the sightings they have had. There is something awe-inspiring about seeing a 25-foot shark approach you, mouth wide open.”
The cruise company has raised concerns about basking sharks, the second biggest fish behind whale sharks, after spotting a number of sharks with chunks missing from their fins. It said the most likely cause for the damage was encounters with boats navigated by negligent skippers and holidaymakers. AK stressed the importance of following the guidelines from the Shark Trust when around the animals.
The swelling numbers led Penzance-based operator Marine Discovery to urge people to be more cautious than normal when on the water around the south west coast.
A spokesman said: “At this time of year basking sharks can be found feeding off the Cornish coast and it’s fantastic to see them. However it is important to remember that they need to be approached carefully so as not to disturb their natural behaviour, this feeding time is a crucial part of their yearly cycle.
“If a shark thrashes its tail and dives or stops feeding and dives then it is likely you have disturbed it. If this happens learn from the mistake and try not to repeat it.”
Exeter University student Tom Whitlock said he saw four basking sharks, one of three plankton-eating sharks alongside the whale and megamouth sharks, on a recent cruise trip along the coast.
Basking sharks became a protected species in 1998 meaning they cannot be targeted, retained or disturbed in British waters.
A spokesperson for the Shark Trust said: “It may come as a surprise to many, but sharks are a natural part of UK marine fauna; whether native or vagrant, over 30 species of shark, as well as over 16 species of skate and ray, can be found in British waters. However, shark, skate and ray numbers have dropped dramatically in our waters due to the impact of poorly managed fisheries.
“Sightings of sharks are mainly reported in summer months when more people are out on the water and should be treated as a privilege rather than a point of concern. Sharks make an easy target for dramatic headlines but it remains far more dangerous to drive to the beach than to swim in our seas.”
British sharks
At least 21 species of shark are resident inhabitants and commonly found around the coasts of Britain all year round, including the Smallspotted Catshark, Porbeagle Shark and Basking Shark, according to The Shark Trust.
Its website says: “Blue Sharks and Shortfin Mako Sharks are seasonal visitors, appearing in British waters in summer during their trans-Atlantic migrations. A few species, Smooth Hammerhead and Frilled Shark may be vagrants, occurring infrequently off the British coast, with their main distribution ranges being outside British waters. At least 11 shark species, including the Portuguese Dogfish, Black Dogfish, Kitefin Shark and Gulper Sharks are only found in deep water.”
|
|
|
|