
Cetorhinus maximus-f
(Gunnerus, 1765)
Classification: Elasmobranchii Lamniformes Cetorhinidae
Reference of the original description
Brugden (Squalus maximus), Beskrvenen ved J. E. Gunnerus. Det Trondhiemske Selskabs Skrifter, 3: 33–49, pl. 2
Brugden (Squalus maximus), Beskrvenen ved J. E. Gunnerus. Det Trondhiemske Selskabs Skrifter, 3: 33–49, pl. 2
Synonyms / new combinations and misspellings
Cetorhinus aff. maximus-f, Cetorhinus cf. maximus-f, Selache maximus-f
Cetorhinus aff. maximus-f, Cetorhinus cf. maximus-f, Selache maximus-f
Types
Cetorhinus maximus-f
Cetorhinus maximus-f
Description:
Citation: Cetorhinus maximus-f (Gunnerus, 1765): In: Database of fossil elasmobranch teeth www.shark-references.com, World Wide Web electronic publication, Version 08/2026
Please send your images of "Cetorhinus maximus-f" to info@shark-references.com
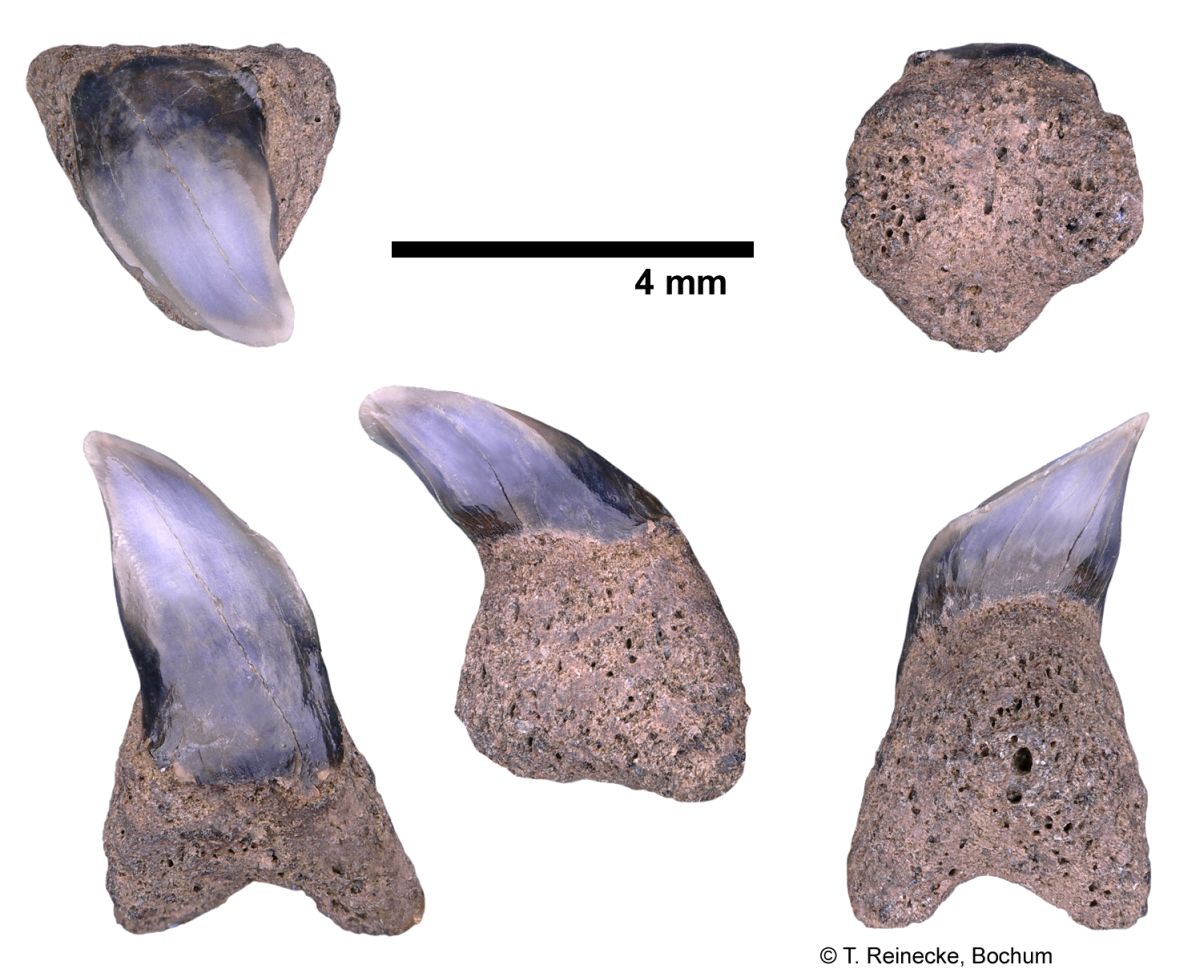
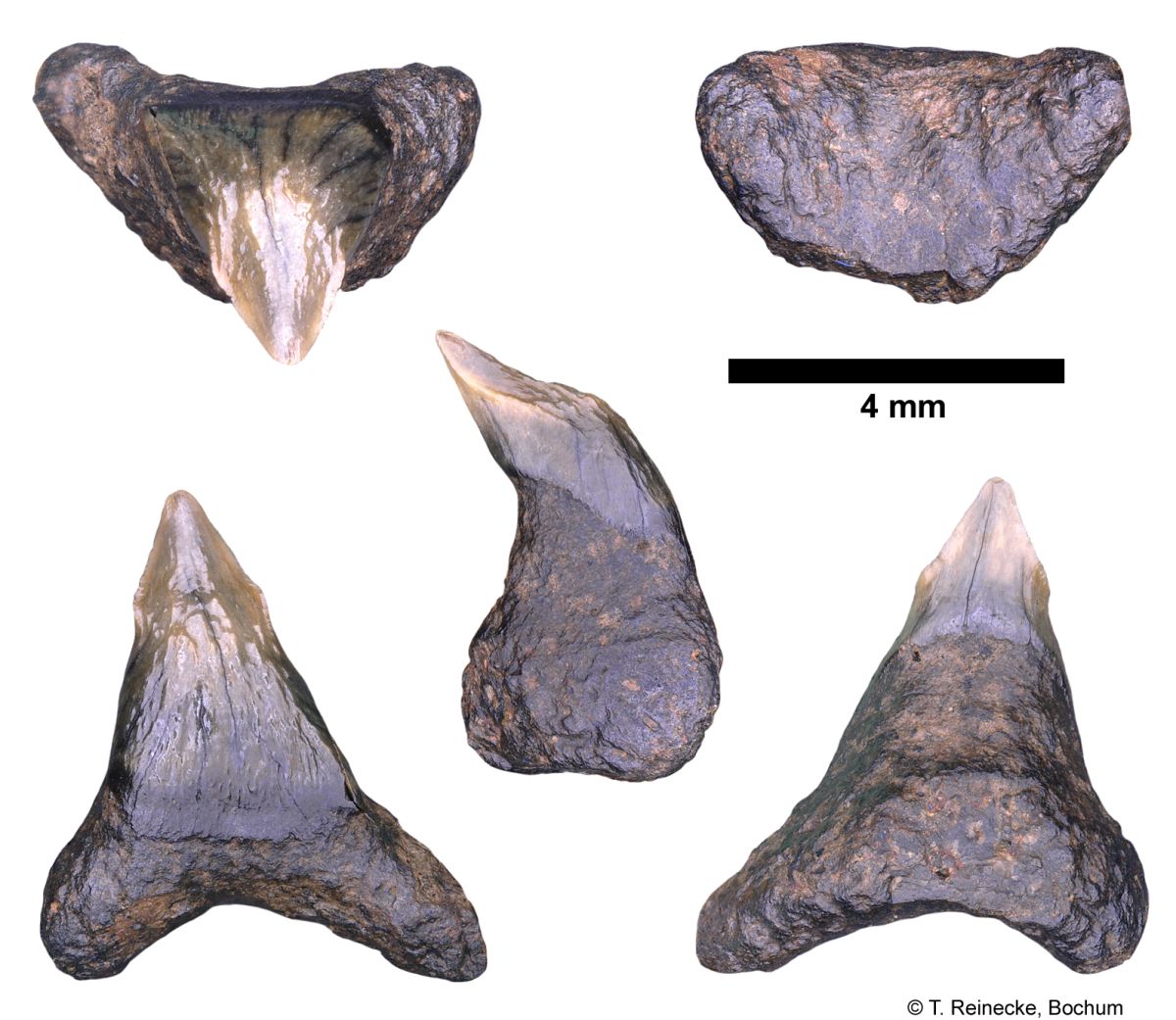
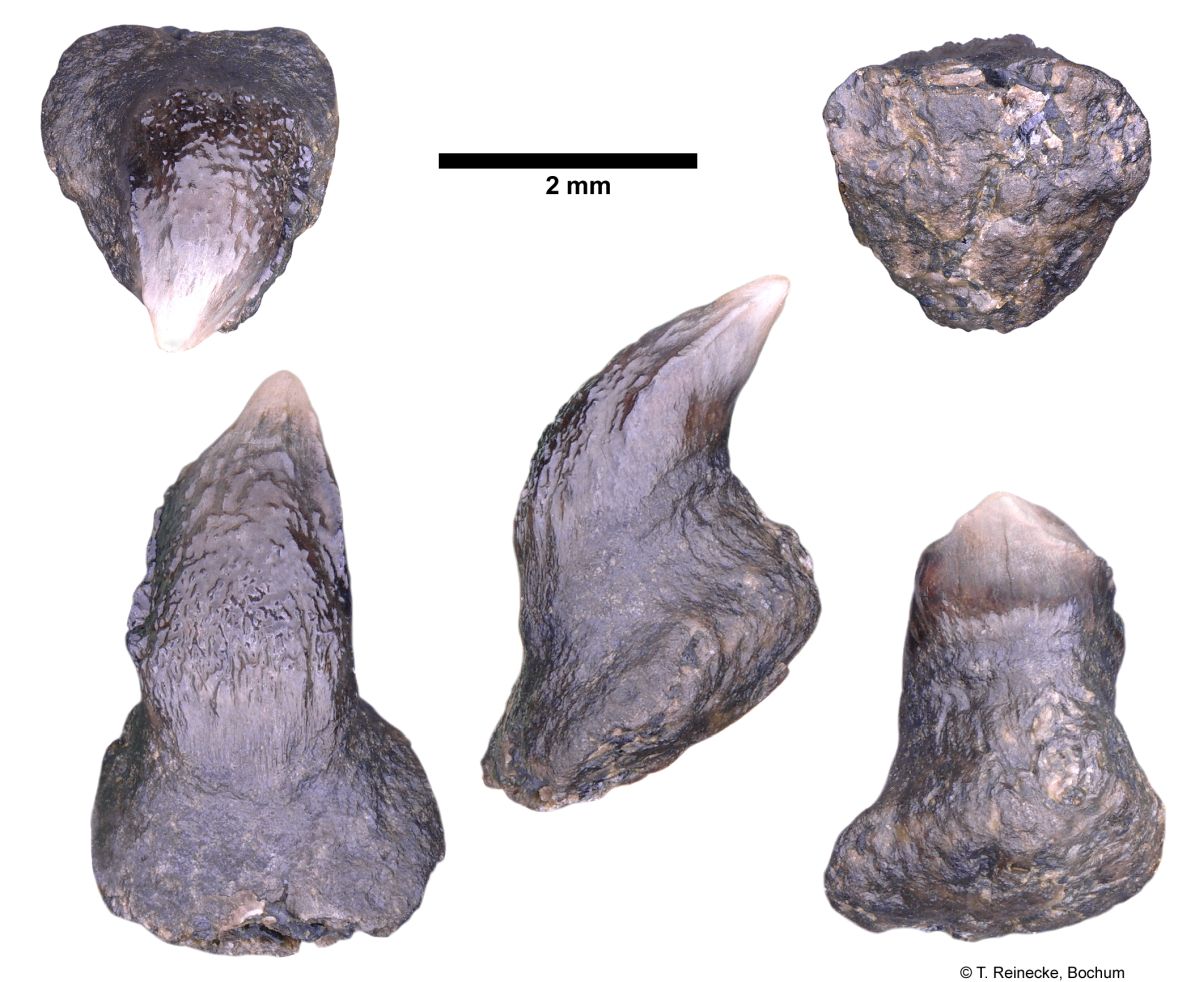
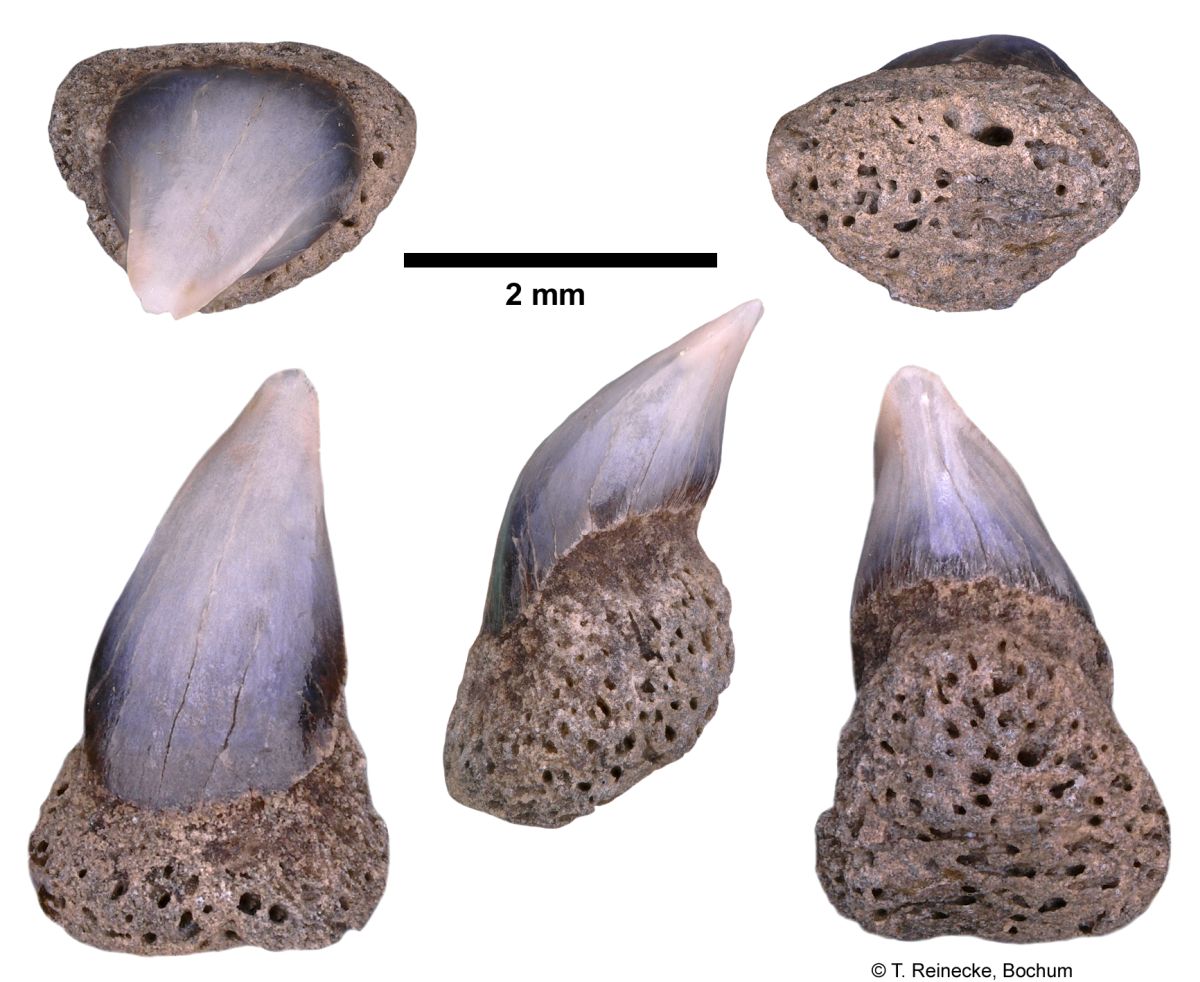
Cetorhinus maximus (Gunner, 1765); teeth, variably worn, collected ex-situ from Breda Formation, early-middle Tortonian, Late Miocene, or Oosterhout Formation, Pliocene; Langenboom, The Netherlands; coll. J. Pollerspöck, Stephansposching, © T. Reinecke, Bochum
Cetorhinus maximus (Gunner, 1765); teeth, variably worn, collected ex-situ from Breda Formation, early-middle Tortonian, Late Miocene, or Oosterhout Formation, Pliocene; Langenboom, The Netherlands; coll. J. Pollerspöck, Stephansposching, © T. Reinecke, Bochum
Description
Description of the Dentition and Gill Rakers of Cetorhinus maximus after Welton (2013) p. 5-22 [17769] (Note: the cited figures can be taken from the original publication (open access):
Dentition
Based on the specimens examined (Figs. 1a-e, 2-9), the dentition of the basking shark Cetorhinus maximus, shows weak gradient monognathic heterodonty in upper and lower jaws (Figs. 6, 7), weak dignathic heterodonty, no evidence of disjunct monognathic heterodonty (no tooth row groups) in either jaw, strong ontogenetic heterodonty in both sexes (Fig. 8), and no dental sexual dimorphism. The occurrence of abnormal teeth, especially in the upper dental series, is relatively common (Fig. 9).
General Tooth Attributes:
Figures 2 through 5 illustrate individual teeth, from selected tooth rows, in the upper and lower dental series of an adult male, and juvenile female Cetorhinus maximus. In all figures, row 1 is the tooth row closest to the jaw symphysis, and the highest numbered row is closest to the distal end of the dental series. All references to the dental series pertain to the total number of tooth rows in the dental band, to the left or right of the jaw symphysis, in the upper or lower jaw. The maximum tooth size for the basking shark is generally < 1 cm, and close to about 6 mm. The crown height is equal or slightly shorter than the root height (in mesial view). The crown is developed into a single, apicobasally compressed, blade-like cusp with convex lingual and labial faces. The cusp is slightly twisted in its vertical axis. Mesial and distal cutting edges are continuous across the rounded cusp apex, and extend basally for about half the crown height in adults, and extend on to mesial and distal blades near the crown foot in juvenile teeth. In labial view, the distal crown foot is constricted, above which it expands into an angular to convex blade whereas the mesial crown base remains straight. The mesial and distal cusp margins are nearly straight. The labial crown foot has numerous, coarse vertical enameloid folds in juvenile teeth, or much reduced folding in adults. These folds originate at the crown foot and extend apically to a level approximately equal to the most apical extent of the lingual neck. These folds grade into irregular, subparallel, to anastomosing, low-relief ridges that extend apically to about half the cusp height. The number, distribution, and strength of these ridges and folds are highly variable. Elsewhere, the enameloid is smooth. A weak labialbasal ledge may be developed, but is usually absent at the crown-neck junction, and is moderately developed lingually. A wide neck underlies the cusp. Its basal contact with the root is nearly flat, and its apical border is widest along the midline of the cusp. Evenlyspaced, deep to shallow, narrow vertical troughs extend across the neck lingually. The cusp that comprises the entire crown is short and elongate – triangular in labial view, and wedge shaped in mesial or distal view. The crown is inclined almost horizontally in juvenile teeth, and is much more erect in adult teeth. The cusp apex extends well beyond the lingual protuberance when viewed mesially or distally in teeth of all age groups. The cusp apex in juvenile teeth is sharp, and is broadly rounded in adult teeth. Cusplets, or cusplet-like projections are never developed, except in some abnormal teeth (see Fig. 9). Labiolingually elongated mesial and distal blades are developed in many teeth of juvenile individuals (Figs. 4- 5). In these teeth, the mesial and distal cutting edges extend the length of the blade, although they may be discontinuous, and a shallow to deep longitudinal trough is present labially, at the junction of the blade and crown. In labial or lingual view, cusps show a weak to moderate increase in distal inclination from row 1 to the distal end of the dental series (Figs. 7, 1a-b, 2a-b). A broad neck is developed at the crown foot lingually, and generally becomes very narrow as it encircles the crown labially. The adult root is massive, nearly circular in mesial view, and more angular, with a labial-basal elongation in juvenile teeth. A weak to strongly convex protuberance occupies the entire lingual face. The basal root border is nearly flat to strongly convex in mesial view. Root lobes are usually absent, but are very rudimentary when developed. Some roots, especially in larger adult teeth, may develop short, blunt, lobate mesial and distal projections midway between the crown foot and root. These projections do not extend lingually, and they may be bordered lingually by a shallow depression. A unique central lingual foramen, or several small, decentralized foramina, may be present on the lingual protuberance. The root is perforated on all surfaces by numerous small foramina. A central transverse groove is rarely present on the lingual protuberance in adult teeth. Single to multiple transverse grooves are more commonly present on the lingual protuberance of juvenile individuals.
Gradient Monognathic Heterodonty:
Gradient monognathic heterodonty is weakly developed in both jaws (Herman et al., 1993) [1136]. In the adult male dental series (LACM 45528-1, Figs. 2, 3, 6-1a-b, 7-1a-b), tooth size increases in the upper and lower dental series from row 1 to about the midpoint of the dental series (rows 49 to 67 in the upper dentition, and rows 5 to 79 in the lower dentition), and then gradually decreases in size to the distal end of the dental band. The cusps are erect at about the same angle across most of the dental series, excluding lower rows 1, 125-129 (Fig. 6-1b), and decrease in height from the middle of the dental series, toward the mesial and distal ends of the dental band (best developed in the upper dental series; see Figure 6-1a). Cutting edges are longest in the largest teeth, and progressively shorten toward the mesial and distal ends of the dental series. Aside from row 1 in the lower dental series, which is a very small tooth with a lingually directed, horizontal crown, all other teeth are slightly more lingually inclined relative to the uppers, and only change significantly in lingual inclination at the distal end of the series in the juvenile dentition (Fig. 6-1b, rows 125-129). In lingual view (Fig. 7), the crowns of mesial rows have weak distal inclination, and this inclination increases distally in the upper dentition, but remains relatively unchanged in the lower tooth series. Four small teeth (Fig. 7-1b, rows 126-129) at the end of the dental series have nearly erect cusps. In mesial view (Fig. 6), the lingual protuberance is very pronounced in the largest teeth, and gradually decreases in convexity toward the mesial and distal-most tooth rows. The dentition of LACM 44280-1 (540 cm (TL) female) shows no significant variation in lingual crown inclination between the upper and lower dentition, or from mesial to distal ends of the dental series (Figs. 6- 2a-b). Distal inclination of the crowns is gradational from near the symphysis, where cusps are almost vertical, to the distal end of the dental series, where cusps have only moderate distal inclination. There is a high degree of variability in the number and occurrence of enameloid folds at the labial crown foot. In lingual view (Fig. 7-2a-b), roots are highly variable in mesodistal width and apicobasal height. Generally, there is a reduction in root mesiodistal width from the center of the dental series to either the mesial or distal end of the dental series. Root height relative to crown height is quite variable, but remains about the same across the dental series. In mesial view, the lingual root protuberance is strongly convex at the center of the dental series and decreases in convexity towards the mesial and distal ends of the dental series (Fig. 6-2a-b).
Dignathic Heterodonty:
Dignathic heterodonty is weak to moderate (Figs. 6-7). The most notable expression being the larger tooth size in the lower dental series relative to teeth in corresponding tooth rows, in the upper dentition (Figs. 6-7). In adult males, the cusps of upper teeth appear to be slightly more erect than corresponding teeth in the lower dentition. Shimada (2002a) [3497] noted that the lower teeth are morphologically less variable than upper teeth. Compagno (2001) [531] reported 203-225/225-230 total tooth rows for both halves of the upper and lower dentition in Cetorhinus maximus. Shimada (2002b: tab. 6) [2175] reported 100?-131 rows for the upper dentition (row count for half the upper dentition) (200?- 262) and 100?-139 (200?- 278 total) for the lower dentition (400?-556 total rows). The two specimens illustrated in this study have the following row counts: LACM 45528-1, 119 rows (upper right), 129 rows (lower left), and LACM 44280-1, 153 rows (upper left), and 139 rows (lower left).
Disjunct Monognathic Heterodonty:
Disjunct monognathic heterodonty is absent in both jaws (Figs. 6, 7). Aside from small, irregularly formed teeth adjacent to the symphysis (upper and lower row 1, Figs. 6, 7), there is no row-group differentiation within the dental series of either jaw. A somewhat variable, but regular gradation in morphology occurs across the dental band. This gradation also occurs across a significant change in tooth size near the distal end of the dental series (Fig. 6-1b, rows 125-129).
Ontogenetic Heterodonty:
Ontogenetic heterodonty is strongly developed in both males and females of Cetorhinus maximus (Fig. 8) (Herman et al., 1993) [1136]. Juvenile teeth (Figs. 6-2a-b, 7-2a-b; Herman et al., 1993, pls. 39-41) [1136], representing sharks having a total length of less than 6 m, have short, broad, triangular crowns with a horizontal lingual inclination relative to the root. Continuous mesial and distal cutting edges cross the crown apex and extend basally and labially, merging with robust, numerous, short complex enameloid folds at the crown foot. Numerous, short, weakly to strongly developed irregular to subparallel ridges cover the basal labial enameloid folds, and extend close to the cusp apex. Weak to strong, sparse enameloid folds extend from the crown foot toward the cusp apex lingually. Roots have a weakly developed lingual protuberance, a pronounced labial basal projection, a flat to weakly convex basal root margin, and development of weak mesial and distal root lobes, especially in the largest teeth in the central portion of the dental series. Transverse grooves are are rarely developed, but when present occur as one or more narrow to broad grooves, originating near the central lingual foramen (or foramen complex), and extend to the basal root border. Adult teeth (Fig. 6-1a-b, 7-1a-b; Herman et al., 1993, pls. 42-44) [1136], representing individuals having a total length exceeding 6 m, differ from juveniles in having longer, much more erect, and narrower crowns (especially in the upper dental series). Mesial and distal cutting edges are short, and extend from the apex basally for a distance less than half the crown height (upper dental series) to two-thirds the crown height (lower dental series). The labial crown foot has relatively few, weakly developed, short enameloid folds or short, irregular subparallel enameloid ridges. The presence of ridges on the labial crown face is highly variable, and when present are sparsely distributed. Roots are massive and bulbous, with a weak to strongly convex lingual protuberance, and flat to strongly convex basal margin. Transverse grooves on the lingual protuberance are rarely developed. Root lobes are absent.
Sexual Heterodonty:
Sexual heterodonty is unknown. The existing sample size is not sufficient to determine the presence or absence of dental sexual dimorphism.
Abnormal Dentition:
Shimada (2002a, fig. 3) [3497] describes and figures a number of abnormal teeth from the upper dentition of Cetorhinus maximus, noting that these teeth are particularly common in the anterior portion of the upper jaw, and especially tooth rows in the symphysial region. Shimada further observes that some of the crowns are reminiscent of the macrophagous lamniforms Alopias Rafinesque, 1810 [1979], Isurus Rafinesque, 1810 [1979], and Pseudocarcharias Cadenat, 1963 [291]. In this study, the upper dentition of LACM 35876-1 also displays a number of anomalous teeth (Fig. 9) with examples of single tooth splitting (Fig. 9, row 30, 31), double splitting (Fig. 9, row 33a) and a trifid split (Fig. 9, row 33b). Two of the three trifid teeth (Fig. 9, rows 33a, 33b) occur in a functional position in the same row, whereas ontogenetically less mature teeth in the same row have normal morphology. A smaller mesodistally compressed tooth (Fig. 9, row 83-84) occurs as an isolated element between normal tooth rows 83 and 84. One instance of row reversal was observed in the upper right jaw, row 1 of LACM 35876-1 (Fig. 7). Several authors (e.g., Pavesi, 1874 [10358]; Compagno, 1990 [520]; Herman, 1979 [1136]; Shimada, 2002a [3497]) describe edentulous spaces and intermittent tooth rows adjacent to the jaw symphysis in the upper and lower jaws; however, the LACM specimens illustrated in Figures 5 and 6, show no break in the dental series. Although the crowns of these aberrant teeth are strikingly different from those of normal Cetorhinus maximus, the roots retain typical attributes of the species. In this regard, isolated abnormal teeth as fossils might be problematic; however, taxonomic confusion arising from their presence can be mitigated by avoiding small sample sizes, in combination with awareness that these anomalies are not unusual in the living species. Underlying this variability is a general morphology that can, in my opinion, be used to characterize the dentition of C. maximus, and separate it from other, as yet undescribed Eocene and younger cetorhinids.
Comments:
Van Den Bosch (1984) [248] described two morphologic tooth types of Recent Cetorhinus. Type 1 teeth are associated with sharks up to 450 cm (TL), and have nearly horizontal crowns with strong labial basal folds in the enameloid at the crown foot. Type 2 teeth are associated with sharks having a total length in the 800 to 1200 cm (TL) range (basking sharks over 1000 cm (TL) are unlikely to exist according to Compagno, 2001, p. 94) [531]. These teeth lack the strong enameloid ornamentation at the labial crown foot, and have relatively smooth, erect crowns. Although Van Den Bosch (1984, p. 214) [248] recognized the possibility that these two tooth types might represent growth stages belonging to one species of Cetorhinus, he ultimately rejected the notion that these differences are ontogenetic. Van Den Bosch’s Type 1 and Type 2 teeth are clearly ontogenetic stages, representing juvenile and adults, respectively. They compare favorably to the ontogenetic series illustrated in Figure 8. Compagno (1990, p. 367) [520] noted that the teeth of Cetorhinus maximus are weakly differentiated into row groups, with a gap between anterior and lateral teeth of the upper jaw. He also stated that tooth roots are “short, high, and flat, with moderately long labial root lobes, small lingual protuberances, and strong basal grooves” (p. 367). I did not find any evidence of row-group differentiation in any of the dentitions examined in this study. Root lobes appear to be more developed in juvenile teeth, and lost in adults. Likewise, basal grooves appear to be an ontogenetic attribute, highly variable in number and distribution, but more commonly present in juvenile teeth, and usually absent in adult teeth. Herman et al. (1993) [1136] figured scanning electron photomicrographs of C. maximus teeth from an 800 cm (TL) female, 386 cm (TL) male, and a 320 cm (TL) male shark. Their observations of tooth morphology, and especially ontogenetic differences, are consistent with those observed in the specimens described here.
Gill Rakers
Van Den Bosch (1984) [248] and Hovestadt and Hovestadt-Euler (2011) [13920] figure gill rakers from Recent Cetorhinus maximus, representing central through distal positions along the gill arch, from individuals of both sexes, and a range of body lengths. In addition to the above, I have illustrated (Fig. 10) five gill rakers, taken from distal to central positions on the gill arch, of a 6.0 to 6.7 m (TL) male C. maximus (LACM 35876- 1). Comparison of C. maximus gill rakers having the same relative position on the gill arch, but from individuals ranging in size from 360 cm (TL) to greater than 600 cm (TL), shows a morphologic (ontogenetic) trend with increasing body length (Hovestadt and Hovestadt-Euler, 2011) [13920]. It is possible that sexual dimorphism, or individual variation, might explain some differences in gill raker morphology; however, the sample size is too small to test this hypothesis. Large adult Cetorhinus maximus gill rakers exceed 20 cm, whereas the largest Keasius parvus, new combination, reaches up to 6 cm (Hovestadt and Hovestadt-Euler, 2011) [13920]. Adult C. maximus gill rakers (6.72 m (TL), Hovestadt and Hovestadt-Euler, 2011, fig. 9, m-u) [13920] have moderately to strongly curved filament bases, and relatively flat to weakly convex distal protuberances. The basal margins are moderately long, and the medial processes are very long and narrow with weakly concave to weakly convex medial edges. Basal widths are narrow, and basal angles are rounded to subangular. Bights are moderately wide and range from angular to subangular. Trends from a distal to central position on the gill arch include increasing curvature of the filament base, increasing basal length, increasing length of the medial process, development of a concave mesial edge of the medial process in the most central gill rakers, and an increase in gill raker length, with the longest gill rakers either in the central-most position on the gill arch (Hovestadt and Hovestadt-Euler, 2011) [13920] or just distal to the central position (LACM 35876-1, Fig. 12-2). The gill rakers of Cetorhinus maximus illustrated in Figure 12 differ from the slightly larger individual described above by having less filament curvature in the most central two positions (Figs. 12-1, 12-2), a base with more convex basal and mesial edges, and a subangular to well rounded basal angle, especially in positions 3 and 4 (Fig. 12). The medial processes are shorter than those in the 672 cm (TL) individual, and, except for the distal most gill raker, they have strongly convex mesial edges (Figs. 12-1 through 12-4). The medial process of the posterior most gill raker (Fig. 12-5) is very short, and superficially resembles the gill rakers of Keasius parvus, new combination, and Keasius taylori, new genus and species. Bight widths are similar, but bight angles are rounded in all but the most distal gill raker, where it is subangular. Gill rakers of a 383-cm (TL) female Cetorhinus maximus (Hovestadt and Hovestadt-Euler, 2011, fig. 9a-h, and a1-f2) [13920] resemble those shown in Figure 12, but differ in the following attributes: the distal half of the series has weakly curved basal filaments, subangular basal angles, flat basal edges, wide bights, and angular to subangular bight angles. The more central gill rakers have a subangular bight, more strongly curved filament bases, and well rounded basal angles, forming a continuous, smoothly curved mesial surface to the base. Ontogenetic changes in the morphology of Cetorhinus maximus gill rakers from a central position on the gill arch, include: an increase in gill raker length, a shift in the basal angle from rounded to subangular, an increase in length of the medial process, and development of a concave mesial edge to the medial process. Variable attributes include degree of filament curvature at its base, basal width, and bight angles that range from curved to angular.
Description of the Dentition and Gill Rakers of Cetorhinus maximus after Welton (2013) p. 5-22 [17769] (Note: the cited figures can be taken from the original publication (open access):
Dentition
Based on the specimens examined (Figs. 1a-e, 2-9), the dentition of the basking shark Cetorhinus maximus, shows weak gradient monognathic heterodonty in upper and lower jaws (Figs. 6, 7), weak dignathic heterodonty, no evidence of disjunct monognathic heterodonty (no tooth row groups) in either jaw, strong ontogenetic heterodonty in both sexes (Fig. 8), and no dental sexual dimorphism. The occurrence of abnormal teeth, especially in the upper dental series, is relatively common (Fig. 9).
General Tooth Attributes:
Figures 2 through 5 illustrate individual teeth, from selected tooth rows, in the upper and lower dental series of an adult male, and juvenile female Cetorhinus maximus. In all figures, row 1 is the tooth row closest to the jaw symphysis, and the highest numbered row is closest to the distal end of the dental series. All references to the dental series pertain to the total number of tooth rows in the dental band, to the left or right of the jaw symphysis, in the upper or lower jaw. The maximum tooth size for the basking shark is generally < 1 cm, and close to about 6 mm. The crown height is equal or slightly shorter than the root height (in mesial view). The crown is developed into a single, apicobasally compressed, blade-like cusp with convex lingual and labial faces. The cusp is slightly twisted in its vertical axis. Mesial and distal cutting edges are continuous across the rounded cusp apex, and extend basally for about half the crown height in adults, and extend on to mesial and distal blades near the crown foot in juvenile teeth. In labial view, the distal crown foot is constricted, above which it expands into an angular to convex blade whereas the mesial crown base remains straight. The mesial and distal cusp margins are nearly straight. The labial crown foot has numerous, coarse vertical enameloid folds in juvenile teeth, or much reduced folding in adults. These folds originate at the crown foot and extend apically to a level approximately equal to the most apical extent of the lingual neck. These folds grade into irregular, subparallel, to anastomosing, low-relief ridges that extend apically to about half the cusp height. The number, distribution, and strength of these ridges and folds are highly variable. Elsewhere, the enameloid is smooth. A weak labialbasal ledge may be developed, but is usually absent at the crown-neck junction, and is moderately developed lingually. A wide neck underlies the cusp. Its basal contact with the root is nearly flat, and its apical border is widest along the midline of the cusp. Evenlyspaced, deep to shallow, narrow vertical troughs extend across the neck lingually. The cusp that comprises the entire crown is short and elongate – triangular in labial view, and wedge shaped in mesial or distal view. The crown is inclined almost horizontally in juvenile teeth, and is much more erect in adult teeth. The cusp apex extends well beyond the lingual protuberance when viewed mesially or distally in teeth of all age groups. The cusp apex in juvenile teeth is sharp, and is broadly rounded in adult teeth. Cusplets, or cusplet-like projections are never developed, except in some abnormal teeth (see Fig. 9). Labiolingually elongated mesial and distal blades are developed in many teeth of juvenile individuals (Figs. 4- 5). In these teeth, the mesial and distal cutting edges extend the length of the blade, although they may be discontinuous, and a shallow to deep longitudinal trough is present labially, at the junction of the blade and crown. In labial or lingual view, cusps show a weak to moderate increase in distal inclination from row 1 to the distal end of the dental series (Figs. 7, 1a-b, 2a-b). A broad neck is developed at the crown foot lingually, and generally becomes very narrow as it encircles the crown labially. The adult root is massive, nearly circular in mesial view, and more angular, with a labial-basal elongation in juvenile teeth. A weak to strongly convex protuberance occupies the entire lingual face. The basal root border is nearly flat to strongly convex in mesial view. Root lobes are usually absent, but are very rudimentary when developed. Some roots, especially in larger adult teeth, may develop short, blunt, lobate mesial and distal projections midway between the crown foot and root. These projections do not extend lingually, and they may be bordered lingually by a shallow depression. A unique central lingual foramen, or several small, decentralized foramina, may be present on the lingual protuberance. The root is perforated on all surfaces by numerous small foramina. A central transverse groove is rarely present on the lingual protuberance in adult teeth. Single to multiple transverse grooves are more commonly present on the lingual protuberance of juvenile individuals.
Gradient Monognathic Heterodonty:
Gradient monognathic heterodonty is weakly developed in both jaws (Herman et al., 1993) [1136]. In the adult male dental series (LACM 45528-1, Figs. 2, 3, 6-1a-b, 7-1a-b), tooth size increases in the upper and lower dental series from row 1 to about the midpoint of the dental series (rows 49 to 67 in the upper dentition, and rows 5 to 79 in the lower dentition), and then gradually decreases in size to the distal end of the dental band. The cusps are erect at about the same angle across most of the dental series, excluding lower rows 1, 125-129 (Fig. 6-1b), and decrease in height from the middle of the dental series, toward the mesial and distal ends of the dental band (best developed in the upper dental series; see Figure 6-1a). Cutting edges are longest in the largest teeth, and progressively shorten toward the mesial and distal ends of the dental series. Aside from row 1 in the lower dental series, which is a very small tooth with a lingually directed, horizontal crown, all other teeth are slightly more lingually inclined relative to the uppers, and only change significantly in lingual inclination at the distal end of the series in the juvenile dentition (Fig. 6-1b, rows 125-129). In lingual view (Fig. 7), the crowns of mesial rows have weak distal inclination, and this inclination increases distally in the upper dentition, but remains relatively unchanged in the lower tooth series. Four small teeth (Fig. 7-1b, rows 126-129) at the end of the dental series have nearly erect cusps. In mesial view (Fig. 6), the lingual protuberance is very pronounced in the largest teeth, and gradually decreases in convexity toward the mesial and distal-most tooth rows. The dentition of LACM 44280-1 (540 cm (TL) female) shows no significant variation in lingual crown inclination between the upper and lower dentition, or from mesial to distal ends of the dental series (Figs. 6- 2a-b). Distal inclination of the crowns is gradational from near the symphysis, where cusps are almost vertical, to the distal end of the dental series, where cusps have only moderate distal inclination. There is a high degree of variability in the number and occurrence of enameloid folds at the labial crown foot. In lingual view (Fig. 7-2a-b), roots are highly variable in mesodistal width and apicobasal height. Generally, there is a reduction in root mesiodistal width from the center of the dental series to either the mesial or distal end of the dental series. Root height relative to crown height is quite variable, but remains about the same across the dental series. In mesial view, the lingual root protuberance is strongly convex at the center of the dental series and decreases in convexity towards the mesial and distal ends of the dental series (Fig. 6-2a-b).
Dignathic Heterodonty:
Dignathic heterodonty is weak to moderate (Figs. 6-7). The most notable expression being the larger tooth size in the lower dental series relative to teeth in corresponding tooth rows, in the upper dentition (Figs. 6-7). In adult males, the cusps of upper teeth appear to be slightly more erect than corresponding teeth in the lower dentition. Shimada (2002a) [3497] noted that the lower teeth are morphologically less variable than upper teeth. Compagno (2001) [531] reported 203-225/225-230 total tooth rows for both halves of the upper and lower dentition in Cetorhinus maximus. Shimada (2002b: tab. 6) [2175] reported 100?-131 rows for the upper dentition (row count for half the upper dentition) (200?- 262) and 100?-139 (200?- 278 total) for the lower dentition (400?-556 total rows). The two specimens illustrated in this study have the following row counts: LACM 45528-1, 119 rows (upper right), 129 rows (lower left), and LACM 44280-1, 153 rows (upper left), and 139 rows (lower left).
Disjunct Monognathic Heterodonty:
Disjunct monognathic heterodonty is absent in both jaws (Figs. 6, 7). Aside from small, irregularly formed teeth adjacent to the symphysis (upper and lower row 1, Figs. 6, 7), there is no row-group differentiation within the dental series of either jaw. A somewhat variable, but regular gradation in morphology occurs across the dental band. This gradation also occurs across a significant change in tooth size near the distal end of the dental series (Fig. 6-1b, rows 125-129).
Ontogenetic Heterodonty:
Ontogenetic heterodonty is strongly developed in both males and females of Cetorhinus maximus (Fig. 8) (Herman et al., 1993) [1136]. Juvenile teeth (Figs. 6-2a-b, 7-2a-b; Herman et al., 1993, pls. 39-41) [1136], representing sharks having a total length of less than 6 m, have short, broad, triangular crowns with a horizontal lingual inclination relative to the root. Continuous mesial and distal cutting edges cross the crown apex and extend basally and labially, merging with robust, numerous, short complex enameloid folds at the crown foot. Numerous, short, weakly to strongly developed irregular to subparallel ridges cover the basal labial enameloid folds, and extend close to the cusp apex. Weak to strong, sparse enameloid folds extend from the crown foot toward the cusp apex lingually. Roots have a weakly developed lingual protuberance, a pronounced labial basal projection, a flat to weakly convex basal root margin, and development of weak mesial and distal root lobes, especially in the largest teeth in the central portion of the dental series. Transverse grooves are are rarely developed, but when present occur as one or more narrow to broad grooves, originating near the central lingual foramen (or foramen complex), and extend to the basal root border. Adult teeth (Fig. 6-1a-b, 7-1a-b; Herman et al., 1993, pls. 42-44) [1136], representing individuals having a total length exceeding 6 m, differ from juveniles in having longer, much more erect, and narrower crowns (especially in the upper dental series). Mesial and distal cutting edges are short, and extend from the apex basally for a distance less than half the crown height (upper dental series) to two-thirds the crown height (lower dental series). The labial crown foot has relatively few, weakly developed, short enameloid folds or short, irregular subparallel enameloid ridges. The presence of ridges on the labial crown face is highly variable, and when present are sparsely distributed. Roots are massive and bulbous, with a weak to strongly convex lingual protuberance, and flat to strongly convex basal margin. Transverse grooves on the lingual protuberance are rarely developed. Root lobes are absent.
Sexual Heterodonty:
Sexual heterodonty is unknown. The existing sample size is not sufficient to determine the presence or absence of dental sexual dimorphism.
Abnormal Dentition:
Shimada (2002a, fig. 3) [3497] describes and figures a number of abnormal teeth from the upper dentition of Cetorhinus maximus, noting that these teeth are particularly common in the anterior portion of the upper jaw, and especially tooth rows in the symphysial region. Shimada further observes that some of the crowns are reminiscent of the macrophagous lamniforms Alopias Rafinesque, 1810 [1979], Isurus Rafinesque, 1810 [1979], and Pseudocarcharias Cadenat, 1963 [291]. In this study, the upper dentition of LACM 35876-1 also displays a number of anomalous teeth (Fig. 9) with examples of single tooth splitting (Fig. 9, row 30, 31), double splitting (Fig. 9, row 33a) and a trifid split (Fig. 9, row 33b). Two of the three trifid teeth (Fig. 9, rows 33a, 33b) occur in a functional position in the same row, whereas ontogenetically less mature teeth in the same row have normal morphology. A smaller mesodistally compressed tooth (Fig. 9, row 83-84) occurs as an isolated element between normal tooth rows 83 and 84. One instance of row reversal was observed in the upper right jaw, row 1 of LACM 35876-1 (Fig. 7). Several authors (e.g., Pavesi, 1874 [10358]; Compagno, 1990 [520]; Herman, 1979 [1136]; Shimada, 2002a [3497]) describe edentulous spaces and intermittent tooth rows adjacent to the jaw symphysis in the upper and lower jaws; however, the LACM specimens illustrated in Figures 5 and 6, show no break in the dental series. Although the crowns of these aberrant teeth are strikingly different from those of normal Cetorhinus maximus, the roots retain typical attributes of the species. In this regard, isolated abnormal teeth as fossils might be problematic; however, taxonomic confusion arising from their presence can be mitigated by avoiding small sample sizes, in combination with awareness that these anomalies are not unusual in the living species. Underlying this variability is a general morphology that can, in my opinion, be used to characterize the dentition of C. maximus, and separate it from other, as yet undescribed Eocene and younger cetorhinids.
Comments:
Van Den Bosch (1984) [248] described two morphologic tooth types of Recent Cetorhinus. Type 1 teeth are associated with sharks up to 450 cm (TL), and have nearly horizontal crowns with strong labial basal folds in the enameloid at the crown foot. Type 2 teeth are associated with sharks having a total length in the 800 to 1200 cm (TL) range (basking sharks over 1000 cm (TL) are unlikely to exist according to Compagno, 2001, p. 94) [531]. These teeth lack the strong enameloid ornamentation at the labial crown foot, and have relatively smooth, erect crowns. Although Van Den Bosch (1984, p. 214) [248] recognized the possibility that these two tooth types might represent growth stages belonging to one species of Cetorhinus, he ultimately rejected the notion that these differences are ontogenetic. Van Den Bosch’s Type 1 and Type 2 teeth are clearly ontogenetic stages, representing juvenile and adults, respectively. They compare favorably to the ontogenetic series illustrated in Figure 8. Compagno (1990, p. 367) [520] noted that the teeth of Cetorhinus maximus are weakly differentiated into row groups, with a gap between anterior and lateral teeth of the upper jaw. He also stated that tooth roots are “short, high, and flat, with moderately long labial root lobes, small lingual protuberances, and strong basal grooves” (p. 367). I did not find any evidence of row-group differentiation in any of the dentitions examined in this study. Root lobes appear to be more developed in juvenile teeth, and lost in adults. Likewise, basal grooves appear to be an ontogenetic attribute, highly variable in number and distribution, but more commonly present in juvenile teeth, and usually absent in adult teeth. Herman et al. (1993) [1136] figured scanning electron photomicrographs of C. maximus teeth from an 800 cm (TL) female, 386 cm (TL) male, and a 320 cm (TL) male shark. Their observations of tooth morphology, and especially ontogenetic differences, are consistent with those observed in the specimens described here.
Gill Rakers
Van Den Bosch (1984) [248] and Hovestadt and Hovestadt-Euler (2011) [13920] figure gill rakers from Recent Cetorhinus maximus, representing central through distal positions along the gill arch, from individuals of both sexes, and a range of body lengths. In addition to the above, I have illustrated (Fig. 10) five gill rakers, taken from distal to central positions on the gill arch, of a 6.0 to 6.7 m (TL) male C. maximus (LACM 35876- 1). Comparison of C. maximus gill rakers having the same relative position on the gill arch, but from individuals ranging in size from 360 cm (TL) to greater than 600 cm (TL), shows a morphologic (ontogenetic) trend with increasing body length (Hovestadt and Hovestadt-Euler, 2011) [13920]. It is possible that sexual dimorphism, or individual variation, might explain some differences in gill raker morphology; however, the sample size is too small to test this hypothesis. Large adult Cetorhinus maximus gill rakers exceed 20 cm, whereas the largest Keasius parvus, new combination, reaches up to 6 cm (Hovestadt and Hovestadt-Euler, 2011) [13920]. Adult C. maximus gill rakers (6.72 m (TL), Hovestadt and Hovestadt-Euler, 2011, fig. 9, m-u) [13920] have moderately to strongly curved filament bases, and relatively flat to weakly convex distal protuberances. The basal margins are moderately long, and the medial processes are very long and narrow with weakly concave to weakly convex medial edges. Basal widths are narrow, and basal angles are rounded to subangular. Bights are moderately wide and range from angular to subangular. Trends from a distal to central position on the gill arch include increasing curvature of the filament base, increasing basal length, increasing length of the medial process, development of a concave mesial edge of the medial process in the most central gill rakers, and an increase in gill raker length, with the longest gill rakers either in the central-most position on the gill arch (Hovestadt and Hovestadt-Euler, 2011) [13920] or just distal to the central position (LACM 35876-1, Fig. 12-2). The gill rakers of Cetorhinus maximus illustrated in Figure 12 differ from the slightly larger individual described above by having less filament curvature in the most central two positions (Figs. 12-1, 12-2), a base with more convex basal and mesial edges, and a subangular to well rounded basal angle, especially in positions 3 and 4 (Fig. 12). The medial processes are shorter than those in the 672 cm (TL) individual, and, except for the distal most gill raker, they have strongly convex mesial edges (Figs. 12-1 through 12-4). The medial process of the posterior most gill raker (Fig. 12-5) is very short, and superficially resembles the gill rakers of Keasius parvus, new combination, and Keasius taylori, new genus and species. Bight widths are similar, but bight angles are rounded in all but the most distal gill raker, where it is subangular. Gill rakers of a 383-cm (TL) female Cetorhinus maximus (Hovestadt and Hovestadt-Euler, 2011, fig. 9a-h, and a1-f2) [13920] resemble those shown in Figure 12, but differ in the following attributes: the distal half of the series has weakly curved basal filaments, subangular basal angles, flat basal edges, wide bights, and angular to subangular bight angles. The more central gill rakers have a subangular bight, more strongly curved filament bases, and well rounded basal angles, forming a continuous, smoothly curved mesial surface to the base. Ontogenetic changes in the morphology of Cetorhinus maximus gill rakers from a central position on the gill arch, include: an increase in gill raker length, a shift in the basal angle from rounded to subangular, an increase in length of the medial process, and development of a concave mesial edge to the medial process. Variable attributes include degree of filament curvature at its base, basal width, and bight angles that range from curved to angular.
Remarks
shark-references Species-ID=16766
shark-references Species-ID=16766
Image gallery














References

Diving into the past: a new assemblage of Neogene elasmobranch microfossils from the eastern Pacific of South America. Papers in Palaeontology, 11(6), Article e70043
DOI: 10.1002/spp2.70043

Paleozoic, Mesozoic and Cenozoic Chondrichthyes from the Japanese Islands. National Museum of Nature and Science Monographs, 56, 1–184
A Review of the Paleobiology of Some Neogene Sharks and the Fossil Records of Extant Shark Species. Diversity, 16(3), Article 147
DOI: 10.3390/d16030147
Body forms of extant lamniform sharks (Elasmobranchii: Lamniformes), and comments on the morphology of the extinct megatooth shark, Otodus megalodon, and the evolution of lamniform thermophysiology. Historical Biology, 35(1), 139–151
DOI: 10.1080/08912963.2021.2025228
Trophic position of Otodus megalodon and great white sharks through time revealed by zinc isotopes. Nature Communications, 13, Article 2980
DOI: 10.1038/s41467-022-30528-9
Remarkable multicuspid teeth in a new elusive skate (Chondrichthyes, Rajiformes) from the Mediterranean Pliocene. Paläontologische Zeitschrift, 95, 117–128
DOI: 10.1007/s12542-020-00542-7
Geology and Paleontology of the Late Miocene Wilson Grove Formation at Bloomfield Quarry, Sonoma County, California. U.S. Geological Survey Scientific Investigations Report 2019–5021, 77 pp.
DOI: 10.3133/sir20195021
The modulating role of traits on the biogeographic dynamics of chondrichthyans from the Neogene to the present. Paleobiology, 44(2), 251–262
DOI: 10.1017/pab.2018.7
Mill-Langenboom (NL) – eine klassische Fundstelle für neogene marine Vertebraten. Der Steinkern, 23, 12–29
Comparative Taphonomy, Taphofacies, and Bonebeds of the Mio-Pliocene Purisima Formation, Central California: Strong Physical Control on Marine Vertebrate Preservation in Shallow Marine Settings. PLoS ONE, 9(3), Article e91419
DOI: 10.1371/journal.pone.0091419
Cetorhinus cf. C. maximus (Gunnerus) (Lamniformes: Cetorhinidae), A Basking Shark from the Late Miocene Empire Formation, Coos Bay, Oregon. Bulletin of the Southern California Academy of Sciences, 112(2),74–92
DOI: 10.3160/0038-3872-112.2.74
Analisi Sistematica, Paleoecologica e Paleobiogeografica della Selaciofauna plio-pleistocenica del Mediterraneo. UNIVERSITÀ DI PISA, Doktorarbeit
Over fossiele resten van Cetorhinus cf. maximus uit het Plioceen van Kallo (België). Grondboor & Hamer, 59(1), 8–12
Haaien en roggen van Mill. Afzettingen WTKG, 23(3), 56–57
A Review of the Analysis of Fish Remains in Chumash Sites. Pacific Coast Archaeological Society Quarterly, 34(1), 25–58
Die Hai- und Rochenfauna aus dem Miozän (Langenfeldium) von Groß Pampau. Der Geschiebesammler, 31(2), 51–113
Der Glimmerton-Aufschluß Groß Pampau (Langenfeldium, Obermiozän), seine Entwicklung und Fossilführung. Der Geschiebesammler, 27(4), 143–183
Eine miozäne Scholle aus der Tongrube in Rhaden bei Lamstedt. Aufschluss, 45, 11–25

Fossiles de Belgique. Dents de Requins et de Raies du Tertiaire de la Belgique. Institut royal des Sciences naturelles de Belgique, 184 pp.
Fossilen uit het 4'te Havendok by Kallo, Belgie. Geologisch Museum Universiteit van Amsterdam: 1–111, 24 pls., 1 tab. [in Dutch]
Haaie- en Roggentanden uit het Tertiair van Belgie. Institut royal des Sciences naturelles de Belgique, 1–171
A deep water elasmobranch fauna from a lower Pliocene outcropping (Northern Italy). In Uyeno, T. & Arai, R. & Taniuchi, T. & Matsuura, K. (Eds) Proceedings of the second international conference on Indo-Pacific fishes. Ichthyological Society of Japan, Tokyo: 133–139
Haaietanden en andere Tertiaire visresten uit Kallo (Belgie) [in Dutch]. GEA, 18(4), 125–160
Réflexions sur la systématique des Galeoidei et sur les affinités du genre Cetorhinus à l'occasion de la découverte d'éléments de la denture d'un exemplaire fossile dans les sables du Kattendijk à Kallo (Pliocène inférieur, Belgique). Annales de la société géologique de Belgique, 102, 357–377
Lithostratigraphical and biostratigraphical subdivision of Tertiary deposits (Oligocene - Pliocene) in the Winterswijk - Almelo region (eastern part of the Netherlands). Scripta Geologica, 29, 1–167
Het profiel van de bouwput onder het eerste kanaaldok nabij Kallo, provincie Oost Vlaanderen, België. Mededelingen Van De Werkgroep Voor Tertiaire En Kwartaire Geologie, 11(4), 173–185
Tertiair van de Kuiperberg bij Ootmarsum. Mededelingen Van De Werkgroep Voor Tertiaire En Kwartaire Geologie, 8(2), 25–45
Fish records from the Pleistocene of southern California in the collections of the Los Angeles County Museum. Bulletin of the Southern California Academy of Sciences, 55, 47–49
Fishes in Upper Miocene and Lower Pleistocene deposits in the Netherlands. Mededeelingen van de Geologische Stichting, Nieuwe Serie, 7, 5–12
Mémoire sur la constitution géognostique de la province de Brabant. Mémoire de la Academie Royal de Belgique, 12(3): 193 p., 4 pl.
Diving into the past: a new assemblage of Neogene elasmobranch microfossils from the eastern Pacific of South America. Papers in Palaeontology, 11(6), Article e70043
DOI: 10.1002/spp2.70043
Paleozoic, Mesozoic and Cenozoic Chondrichthyes from the Japanese Islands. National Museum of Nature and Science Monographs, 56, 1–184
A Review of the Paleobiology of Some Neogene Sharks and the Fossil Records of Extant Shark Species. Diversity, 16(3), Article 147
DOI: 10.3390/d16030147
Body forms of extant lamniform sharks (Elasmobranchii: Lamniformes), and comments on the morphology of the extinct megatooth shark, Otodus megalodon, and the evolution of lamniform thermophysiology. Historical Biology, 35(1), 139–151
DOI: 10.1080/08912963.2021.2025228
Trophic position of Otodus megalodon and great white sharks through time revealed by zinc isotopes. Nature Communications, 13, Article 2980
DOI: 10.1038/s41467-022-30528-9
Remarkable multicuspid teeth in a new elusive skate (Chondrichthyes, Rajiformes) from the Mediterranean Pliocene. Paläontologische Zeitschrift, 95, 117–128
DOI: 10.1007/s12542-020-00542-7
Geology and Paleontology of the Late Miocene Wilson Grove Formation at Bloomfield Quarry, Sonoma County, California. U.S. Geological Survey Scientific Investigations Report 2019–5021, 77 pp.
DOI: 10.3133/sir20195021
The modulating role of traits on the biogeographic dynamics of chondrichthyans from the Neogene to the present. Paleobiology, 44(2), 251–262
DOI: 10.1017/pab.2018.7
Mill-Langenboom (NL) – eine klassische Fundstelle für neogene marine Vertebraten. Der Steinkern, 23, 12–29
Comparative Taphonomy, Taphofacies, and Bonebeds of the Mio-Pliocene Purisima Formation, Central California: Strong Physical Control on Marine Vertebrate Preservation in Shallow Marine Settings. PLoS ONE, 9(3), Article e91419
DOI: 10.1371/journal.pone.0091419
Cetorhinus cf. C. maximus (Gunnerus) (Lamniformes: Cetorhinidae), A Basking Shark from the Late Miocene Empire Formation, Coos Bay, Oregon. Bulletin of the Southern California Academy of Sciences, 112(2),74–92
DOI: 10.3160/0038-3872-112.2.74
Analisi Sistematica, Paleoecologica e Paleobiogeografica della Selaciofauna plio-pleistocenica del Mediterraneo. UNIVERSITÀ DI PISA, Doktorarbeit
Over fossiele resten van Cetorhinus cf. maximus uit het Plioceen van Kallo (België). Grondboor & Hamer, 59(1), 8–12
Haaien en roggen van Mill. Afzettingen WTKG, 23(3), 56–57
A Review of the Analysis of Fish Remains in Chumash Sites. Pacific Coast Archaeological Society Quarterly, 34(1), 25–58
Die Hai- und Rochenfauna aus dem Miozän (Langenfeldium) von Groß Pampau. Der Geschiebesammler, 31(2), 51–113
Der Glimmerton-Aufschluß Groß Pampau (Langenfeldium, Obermiozän), seine Entwicklung und Fossilführung. Der Geschiebesammler, 27(4), 143–183
Eine miozäne Scholle aus der Tongrube in Rhaden bei Lamstedt. Aufschluss, 45, 11–25
Fossiles de Belgique. Dents de Requins et de Raies du Tertiaire de la Belgique. Institut royal des Sciences naturelles de Belgique, 184 pp.
Fossilen uit het 4'te Havendok by Kallo, Belgie. Geologisch Museum Universiteit van Amsterdam: 1–111, 24 pls., 1 tab. [in Dutch]
Haaie- en Roggentanden uit het Tertiair van Belgie. Institut royal des Sciences naturelles de Belgique, 1–171
A deep water elasmobranch fauna from a lower Pliocene outcropping (Northern Italy). In Uyeno, T. & Arai, R. & Taniuchi, T. & Matsuura, K. (Eds) Proceedings of the second international conference on Indo-Pacific fishes. Ichthyological Society of Japan, Tokyo: 133–139
Haaietanden en andere Tertiaire visresten uit Kallo (Belgie) [in Dutch]. GEA, 18(4), 125–160
Réflexions sur la systématique des Galeoidei et sur les affinités du genre Cetorhinus à l'occasion de la découverte d'éléments de la denture d'un exemplaire fossile dans les sables du Kattendijk à Kallo (Pliocène inférieur, Belgique). Annales de la société géologique de Belgique, 102, 357–377
Lithostratigraphical and biostratigraphical subdivision of Tertiary deposits (Oligocene - Pliocene) in the Winterswijk - Almelo region (eastern part of the Netherlands). Scripta Geologica, 29, 1–167
Het profiel van de bouwput onder het eerste kanaaldok nabij Kallo, provincie Oost Vlaanderen, België. Mededelingen Van De Werkgroep Voor Tertiaire En Kwartaire Geologie, 11(4), 173–185
Tertiair van de Kuiperberg bij Ootmarsum. Mededelingen Van De Werkgroep Voor Tertiaire En Kwartaire Geologie, 8(2), 25–45
Fish records from the Pleistocene of southern California in the collections of the Los Angeles County Museum. Bulletin of the Southern California Academy of Sciences, 55, 47–49
Fishes in Upper Miocene and Lower Pleistocene deposits in the Netherlands. Mededeelingen van de Geologische Stichting, Nieuwe Serie, 7, 5–12
Mémoire sur la constitution géognostique de la province de Brabant. Mémoire de la Academie Royal de Belgique, 12(3): 193 p., 4 pl.