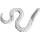

Palaeoxyris humblei
Fischer, Axsmith, & Ash, 2010
Classification: Elasmobranchii
Reference of the original description
First unequivocal record of the hybodont shark egg capsule Palaeoxyris in the Mesozoic of North America. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 255(3), 327–344
First unequivocal record of the hybodont shark egg capsule Palaeoxyris in the Mesozoic of North America. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 255(3), 327–344
Images of types



Description:
Citation: Palaeoxyris humblei Fischer, Axsmith, & Ash, 2010: In: Database of fossil elasmobranch teeth www.shark-references.com, World Wide Web electronic publication, Version 07/2026
Please send your images of "Palaeoxyris humblei" to info@shark-references.com
Palaeoxyris humblei Fischer, Axsmith, & Ash, 2010 © Jan Fischer, Germany
Palaeoxyris humblei Fischer, Axsmith, & Ash, 2010 © Jan Fischer, Germany
Distribution Stratigraphy
Mudstone facies in the Newspaper Rock Bed of the Blue Mesa Member of the Chinle Formation; Late Triassic – Norian (Zone of Dinophyton, Adamanian land mammal faunalchron) [7072];
Mudstone facies in the Newspaper Rock Bed of the Blue Mesa Member of the Chinle Formation; Late Triassic – Norian (Zone of Dinophyton, Adamanian land mammal faunalchron) [7072];
Description
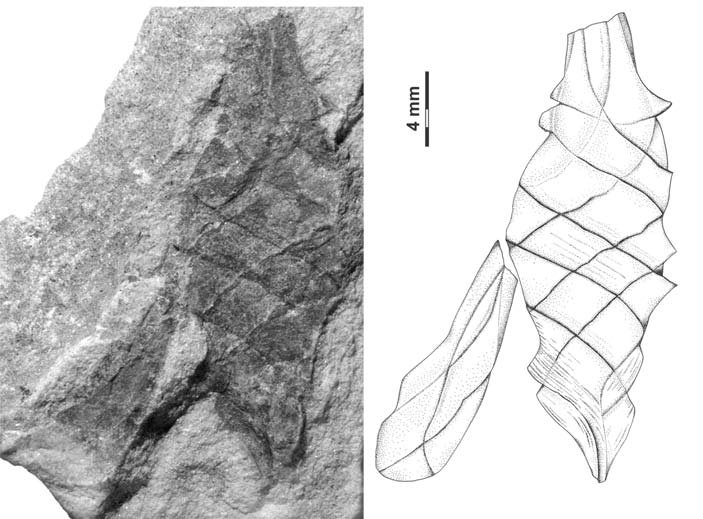
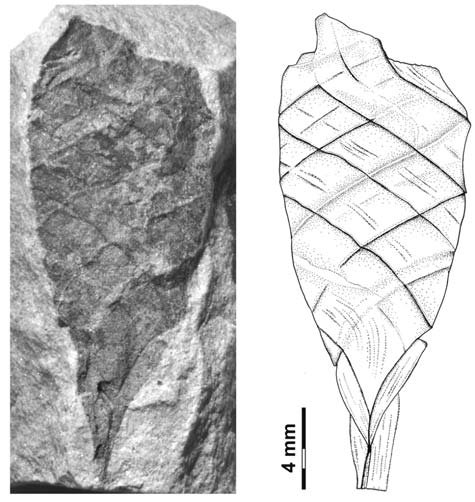
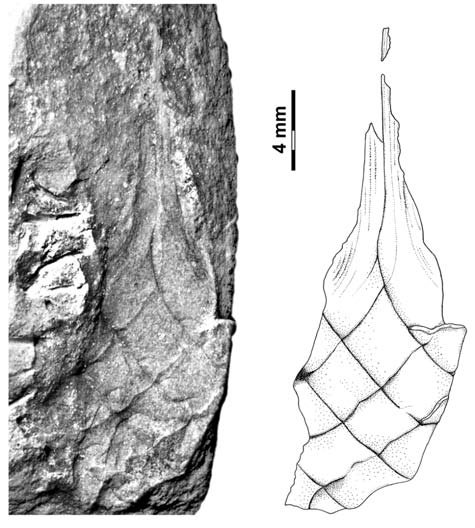
Body broadly fusiform, up to 13.0 mm long, 9.0 mm wide at the widest point, gradually tapering toward each end, traversed by seven segments of three-four helicoidally twisted bands, bands averaging 3.0 mm in width, segments arising at an angle of 20-36º in the middle of the body, 40-50º near the ends, no alternating band pattern; beak at least 12.5 mm long, 0.75 mm wide at the most distally preserved end; pedicle with gradual constriction, ~ 4.6 mm wide at transition point to body, 1.4 mm wide distally, central needle-like structure; bands lined with collarette 1.2 mm in height, distinct fine parallel striation on bands and collarettes [7072];
Body broadly fusiform, up to 13.0 mm long, 9.0 mm wide at the widest point, gradually tapering toward each end, traversed by seven segments of three-four helicoidally twisted bands, bands averaging 3.0 mm in width, segments arising at an angle of 20-36º in the middle of the body, 40-50º near the ends, no alternating band pattern; beak at least 12.5 mm long, 0.75 mm wide at the most distally preserved end; pedicle with gradual constriction, ~ 4.6 mm wide at transition point to body, 1.4 mm wide distally, central needle-like structure; bands lined with collarette 1.2 mm in height, distinct fine parallel striation on bands and collarettes [7072];
Discussion
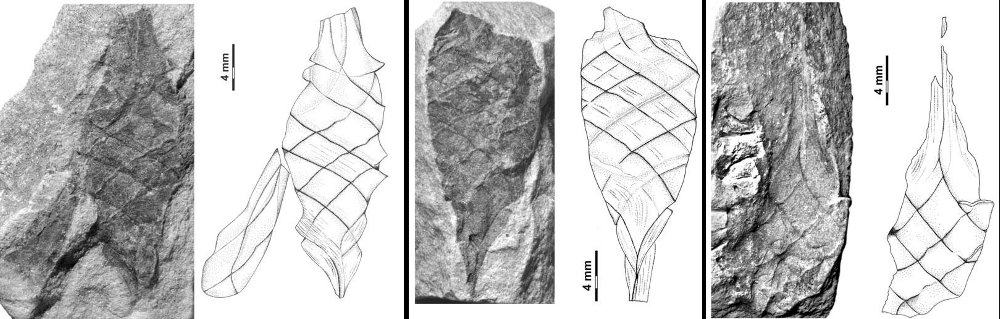
All the specimens described here represent the same type of egg capsule, and based on their size, shape, and banding characteristic are unquestionably assignable to the form genus Palaeoxyris BRONGNIART, 1828. Although all of the specimens are incomplete (Fig. 6), in combination the features of each allow a reasonable reconstruction of the entire capsule (Figs. 6-7). The body/beak length ratio in these specimens averages about 1/1, which is a common value for Palaeoxyris according to MÜLLER (1978). Only a short length of the pedicle is preserved on any of the specimens in our collection, but in comparison to complete specimens of Palaeoxyris (e.g., CROOKALL 1930; MÜLLER 1978) a total body/pedicle length ratio up to 2 seems to be plausible. Furthermore, preservation in specimen PEFO 34350 (Fig. 3) and especially PEFO 34351 (Fig. 4) suggests a subparallel arrangement of the ribs on the pedicle as shown in Fig. 2 similar to other Triassic forms (e.g., BRONGNIART 1828; PRESL 1838; FISCHER et al. 2007) but not to Paleozoic forms (FISCHER et al. 2008b). The body of the new species is traversed by three-four bands. As previously reported by CROOKALL (1928b, 1930) and MÜLLER (1978) there can be a small variation in band numbers within one genus. Therefore, the use of the band number by itself as a diagnostic feature among species of the genus seems to be inadequate without using other parameters. The angle of the bands varies between specimens of a species because of taphonomical modification. The most remarkable feature of the new Chinle fossil compared to other species of Palaeoxyris is the presence of an unusually well-preserved collarette in combination with fine longitudinal striations on the bands (Figs. 2-3). The presence of the collarette has normally been regarded as one of the chief characters distinguishing Palaeoxyris from the morphologically similar egg capsule Fayolia by RENAULLT & ZEILLER (1884) and by CROOKALL (1928a), and facili - tates comparison with modern heterodont shark egg cap - sules (RENAULT & ZEILLER 1888; ZEILLER 1890). Furthermore, similar features have been described in certain other Palaeoxyris specimens. For example, LESQUEREUX (1870, 1880) described and illustrated noticeable pointed, trian - gular “tooth-like” structures (= fragments of the collarette) on the body margin of P. appendiculata from the West - phalian D Mazon Creek locality of Illinois, U.S.A. Such structures were also illustrated by RENAULT & ZEILLER (1888), SAPORTA (1891) and BROWN (1950). In addition, KIDSTON (1886) illustrated similar structures in P. trispiralis from the Westphalian B of Staffordshire, England. Also, parallel striae on their bands were described and figured by LESQUEREUX (1870) in P. appendiculata as well as P. prendeli from the Mazon Creek locality (the later specimen should be revised to P. helicteroides because of the typical alternating band widths) and by SAPORTA (1891) for P. carbonaria. Nevertheless, the excellent preservation of the collarettes on large parts of the P. humblei specimens, especially in the body-pedicle transition (Fig. 3), is not known in any previous records of Palaeoxyris. In fact, it strongly resembles the collarette and the striated capsule surface of modern heterodont egg capsules (see ZIDEK 1976; FISCHER et. 2008a) and supports the assignment of these specimens of Palaeoxyris to the egg capsules of the elasmobranchs [7072];
All the specimens described here represent the same type of egg capsule, and based on their size, shape, and banding characteristic are unquestionably assignable to the form genus Palaeoxyris BRONGNIART, 1828. Although all of the specimens are incomplete (Fig. 6), in combination the features of each allow a reasonable reconstruction of the entire capsule (Figs. 6-7). The body/beak length ratio in these specimens averages about 1/1, which is a common value for Palaeoxyris according to MÜLLER (1978). Only a short length of the pedicle is preserved on any of the specimens in our collection, but in comparison to complete specimens of Palaeoxyris (e.g., CROOKALL 1930; MÜLLER 1978) a total body/pedicle length ratio up to 2 seems to be plausible. Furthermore, preservation in specimen PEFO 34350 (Fig. 3) and especially PEFO 34351 (Fig. 4) suggests a subparallel arrangement of the ribs on the pedicle as shown in Fig. 2 similar to other Triassic forms (e.g., BRONGNIART 1828; PRESL 1838; FISCHER et al. 2007) but not to Paleozoic forms (FISCHER et al. 2008b). The body of the new species is traversed by three-four bands. As previously reported by CROOKALL (1928b, 1930) and MÜLLER (1978) there can be a small variation in band numbers within one genus. Therefore, the use of the band number by itself as a diagnostic feature among species of the genus seems to be inadequate without using other parameters. The angle of the bands varies between specimens of a species because of taphonomical modification. The most remarkable feature of the new Chinle fossil compared to other species of Palaeoxyris is the presence of an unusually well-preserved collarette in combination with fine longitudinal striations on the bands (Figs. 2-3). The presence of the collarette has normally been regarded as one of the chief characters distinguishing Palaeoxyris from the morphologically similar egg capsule Fayolia by RENAULLT & ZEILLER (1884) and by CROOKALL (1928a), and facili - tates comparison with modern heterodont shark egg cap - sules (RENAULT & ZEILLER 1888; ZEILLER 1890). Furthermore, similar features have been described in certain other Palaeoxyris specimens. For example, LESQUEREUX (1870, 1880) described and illustrated noticeable pointed, trian - gular “tooth-like” structures (= fragments of the collarette) on the body margin of P. appendiculata from the West - phalian D Mazon Creek locality of Illinois, U.S.A. Such structures were also illustrated by RENAULT & ZEILLER (1888), SAPORTA (1891) and BROWN (1950). In addition, KIDSTON (1886) illustrated similar structures in P. trispiralis from the Westphalian B of Staffordshire, England. Also, parallel striae on their bands were described and figured by LESQUEREUX (1870) in P. appendiculata as well as P. prendeli from the Mazon Creek locality (the later specimen should be revised to P. helicteroides because of the typical alternating band widths) and by SAPORTA (1891) for P. carbonaria. Nevertheless, the excellent preservation of the collarettes on large parts of the P. humblei specimens, especially in the body-pedicle transition (Fig. 3), is not known in any previous records of Palaeoxyris. In fact, it strongly resembles the collarette and the striated capsule surface of modern heterodont egg capsules (see ZIDEK 1976; FISCHER et. 2008a) and supports the assignment of these specimens of Palaeoxyris to the egg capsules of the elasmobranchs [7072];
Remarks
shark-references Species-ID=8540;
shark-references Species-ID=8540;
References

First unequivocal record of the hybodont shark egg capsule Palaeoxyris in the Mesozoic of North America. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 255(3), 327–344
DOI: 10.1127/0077-7749/2009/0028
First unequivocal record of the hybodont shark egg capsule Palaeoxyris in the Mesozoic of North America. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 255(3), 327–344
DOI: 10.1127/0077-7749/2009/0028