NEWSLETTER 03/2014 18.03.2014
|
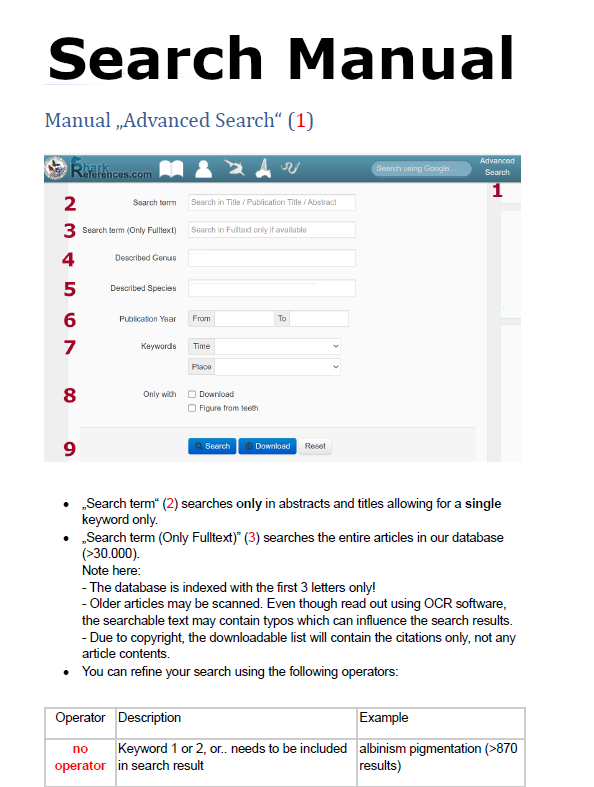
|
|
Please acknowledge use of the database www.shark-references.com in your publications, and cite:
Pollerspöck, J. 2014, Bibliography database of living/fossil sharks, rays and chimaeras (Chondrichtyes: Elasmobranchii, Holocephali), www.shark-references.com, World Wide Web electronic publication, Version 2014 |
|
|
PARTNERS OF SHARK-REFERENCES:
Would you like to become a shark-reference partner? Please contanct me per E-mail!
Partner in Google-Maps: 
|
|
New images at shark-references:
Many thanks to the following persons for the permission to use their images:
- Simon Weigmann, Hamburg for a image of Indobatis ori (WALLACE, 1967):

- William T. White and Peter Last, CSIRO Marine & Atmospheric Research, Wealth from Oceans Flagship, Hobart, Tasmania, Australia for permission to use the images of the CSIRO Marine and Atmospheric Research Papers at shark-references.com, e.g. Lamiopsis tephrodes (FOWLER, 1905):

- João Paulo C. B. Da Silva, Universidade de São Paulo for the images of Potamotrygon limai FONTENELLE, DA SILVA & DE CARVALHO, 2014:

- Mark Sabaj Pérez, for images of some Potamotrygon/Paratrygon species:

see more:
https://shark-references.com/species/view/Potamotrygon-motoro
https://shark-references.com/species/view/Paratrygon-aiereba
https://shark-references.com/species/view/Potamotrygon-orbignyi
- Jacques Herman & Hilde Van Waesacques, Beigem for the image of Pliotrema warreni REGAN, 1906;

- Lawrence R. Frank and Rachel Berquist, Radiology - Center for Scientific Computation in Imaging, University of California San Diego for the image of e.g. Squatina californica AYRES, 1859:

For more images see:
https://shark-references.com/species/view/Hexanchus-griseushttps://shark-references.com/species/view/Pristiophorus-japonicus
Please visit the website of the Digital Fish Library (DFL)
Please support shark-references and send your images to: info@shark-references.com |
|
Missing papers:
Many thanks to all friends of shark-references, who send me some missing papers last month!
Shark-References would kindly like to ask you for your contribution to this project.
At the moment I search the following papers:
SMITH, J.L.B. 1936 Two interesting new fishes from South Africa. Transactions of the Royal Society of South Africa, 24 (1): 1-6, Pls. 1-2
Talwar, P.K. 1981 Identity of the type specimen of the Scyliorhinid Shark Scyllium
hispidum Alcock. Bulletin of Zoological Survey of India 4, 231–234.
Talwar, P.K. 1990 Fishes of Andaman and Nicobar Islands: a synoptic survey. Journal of
the Andaman Science Association Andaman Science Association, Port Blair 6, 71–102.
Please support www.shark-references.com and send missing papers (not listed papers or papers without the infosymbol) to juergen.pollerspoeck@shark-references.com.
|
|
New Books/Papers from Parnter:
Géominpal Belgica
Découvertes géologiques, minéralogiques et paléontologiques
en Belgique. Vol 6
Observations concerning the Evolution and the Parasystematics of all the living and fossil Chlamydoselachiformes, Squatiniformes, Orectolobiformes, and Pristiophoriformes, based on both biological and odontological data. Suggestion of a possible origin of the Order Pristiophoriformes, of the Order Ganopristiformes and a global Synthesis of the previous Systematics proposals. (Last Data integrated: These published before 05.03.2014)
By Herman Jacques & Hilde Van Waes
free download: http://www.geominpal.be/Geominpal%20Belgica%206/Geominpal%20Belgica%206.pdf
example plates:
 
Plate 15 (Echinorhinus brucus (BONNATERRE, 1788) and Plate 16 (Echinorhinus cookei PIETSCHMANN, 1928) |
|
Meetings:

The KwaZulu-Natal Sharks Board is pleased to confirm that the second Sharks International symposium will be held in Durban, South Africa on 2-6 June 2014. This is a sequel to the highly successful inaugural Sharks International meeting in Cairns, Australia in June 2010. The meeting will comprise four full days of presentations, commencing Monday 2 June and ending Friday 6 June, with Wednesday 4 June set aside for a variety of exciting mid-conference excursions.
The aim of this conference is to provide a forum for the world’s leading shark and ray researchers, along with students and early career scientists to meet, share ideas, update information and report on the progress of their most recent scientific studies. We would encourage any researchers and students with a general interest in the marine environment to also attend as networking opportunities at a conference of this nature and size are enormous.
For further information please visit the Symposium’s web page at:
http://www.sharksinternational.org
Any queries regarding the symposium can also be sent to the organising committee at:
sharksinternational2014@gmail.com |
|

IV Encuentro Colombiano sobre Condrictios:
place: Universidad EAFIT de Medellín, Colombia
date: 20. - 24. October 2014
more information
|
|
New described species/Taxonomic News:
Recent:
LOBODA, T.S. & DE CARVALHO, M.R. (2013): Systematic revision of the Potamotrygon motoro (Müller & Henle, 1841) species complex in the Paraná-Paraguay basin, with description of two new ocellated species (Chondrichthyes: Myliobatiformes: Potamotrygonidae). Neotropical Ichthyology, 11 (4): 693-737
New species: Potamotrygon pantanensis, Potamotrygon amandae
Abstract: A systematic revision of the Potamotrygon motoro (Müller & Henle, 1841) species complex in the Paraná-Paraguay basin based on morphological characters was undertaken. Morphological systems analyzed include external morphology, coloration, dermal denticles, and spines, canals of the ventral lateral-line system, and skeletal components. Potamotrygon motoro is widely distributed in the Paraná-Paraguay basin and some of its diagnostic characters are: ocelli present on dorsal disc tricolored, well-defined and evenly distributed, with diameter similar or greater than eye-length; ventral coloration with relatively large whitish central region, with gray or brown area predominant on outer ventral disc margins; dermal denticles well-developed and star-shaped over central disc; labial grooves absent; monognathic heterodonty present in upper and lower jaws of adults. Potamotrygon pauckei Castex, 1963 and Potamotrygon labradori Castex, Maciel & Achenbach, 1963, are synonymized with P. motoro; Potamotrygon alba Castex, 1963, is a nomen dubium in accordance with previous authors. Additionally, two new ocellated species of Potamotrygon from the Paraná-Paraguay basin are described: Potamotrygon pantanensis, sp. nov. and Potamotrygon amandae, sp. nov. These are described and compared with P. motoro and other congeners. Potamotrygon pantanensis, sp. nov. is described from the northern Pantanal region; Potamotrygon amandae, sp. nov. is widespread in the Paraná-Paraguay basin.
WEIGMANN, S. & STEHMANN, M. & THIEL, R. (2014): Complementary redescription of Anacanthobatis ori (Wallace, 1967) and its assignment to Indobatis n. g. (Elasmobranchii, Anacanthobatidae), with comments on other legskates. Zootaxa, 3779 (2): 101-132
New genus: Indobatis
Abstract: Anacanthobatis ori is one of the least known species of the family Anacanthobatidae with only four juvenile specimens reported. The species remained assigned to the genus Anacanthobatis sensu lato due to the lack of an adult male as external and skeletal clasper characters are the essential diagnostic features for the differentiation of genera and subgenera within the family Anacanthobatidae. Since an adult male of A. ori became available, along with an adult female and six further juveniles, the authors reinvestigated the species and present its so far unknown diagnostic characters of clasper morphology and skeleton and scapulocoracoid. The clasper turned out to be the most complex one of all known anacanthobatids as the external components flag, slit, pseudosiphon-like cavity, pecten, and two sentinas are not known from any other anacanthobatid species. Furthermore, a dorsal terminal 1 cartilage is present but displaced proximally of the terminal clasper skeleton, the outer edge of dorsal terminal 2 is deeply serrated, the ventral terminal has a very long, curved, straplike process, and the proximal part of accessory terminal 1 is embedded in the cavity of the baseball-glove-like head of accessory terminal 2. Due to the strong differences in external and internal clasper characters to all other known anacanthobatid species, A. ori is placed in its own, newly erected genus, Indobatis.
Parasitology:
CAIRA, J.N. & JENSEN, K. & WAESCHENBACH, A. & OLSON, P.D. & LITTLEWOOD, D.T.J. (2014): Orders out of chaos - molecular phylogenetics reveals the complexity of shark and stingray tapeworm relationships. International Journal for Parasitology, 44 (1): 55-73
New order: Onchoproteocephalidea, Phyllobothriidea
Abstract: Novel molecular data are presented to resolve the long-standing issue of the non-monophyly of the elasmobranch-hosted tapeworm order Tetraphyllidea relative to the other acetabulate eucestode orders. Bayesian inference analyses of various combinations of full ssrDNA, and full or partial lsrDNA (D1-D3), sequence data, which included 134 species representing 97 genera across the 15 eucestode orders, were conducted. New ssrDNA data were generated for 82 species, partial lsrDNA data for 53 species, and full lsrDNA data for 29 species. The monophyly of each of the elasmobranch-hosted orders Cathetocephalidea, Litobothriidea, Lecanicephalidea and Rhinebothriidea was confirmed, as was the non-monophyly of the Tetraphyllidea. Two relatively stable groups of tetraphyllidean taxa emerged and are hereby designated as new orders. The Onchoproteocephalidea n. ord. is established to recognise the integrated nature of one undescribed and 10 described genera of hook-bearing tetraphyllideans, previously placed in the family Onchobothriidae, with the members of the order Proteocephalidea. The Phyllobothriidea n. ord. is established for a subset of 12 non-hooked genera characterised by scoleces bearing four bothridia each with an anterior accessory sucker; most parasitise sharks and have been assigned to the Phyllobothriidae at one time or another. Tentative ordinal placements are suggested for eight additional genera; placements for the remaining tetraphyllidean genera have not yet emerged. We propose that these 17 genera remain in the "Tetraphyllidea". Among these, particularly labile across analyses were Anthobothrium, Megalonchos, Carpobothrium, Calliobothrium and Caulobothrium. The unique association of Chimaerocestus with holocephalans, rather than with elasmobranchs, appears to represent a host-switching event. Both of the non-elasmobranch hosted clades of acetabulate cestodes (i.e. Proteocephalidea and Cyclophyllidea and their kin) appear to have had their origins with elasmobranch cestodes. Across analyses, the sister group to the clade of "terrestrial" cestode orders was found to be an elasmobranch-hosted genus, as was the sister to the freshwater fish- and tetrapod-hosted Proteocephalidea. Whilst further data are required to resolve outstanding nomenclatural and phylogenetic issues, the present analyses contribute significantly to an understanding of the evolutionary radiation of the entire Cestoda. Clearly, elasmobranch tapeworms comprise the backbone of cestode phylogeny.
Fossil:
SHIMADA, K. & WELTON, B.J. & LONG, D.J. (2014): A new fossil megamouth shark (Lamniformes, Megachasmidae) from the Oligocene-Miocene of the western United States. Journal of Vertebrate Paleontology, 34 (2): 281-290
New species: Megachasma applegatei
Abstract: The extant megamouth shark, Megachasma pelagios (Lamniformes: Megachasmidae), is a large filter-feeding fish. We here describe a new species of Megachasma, M. applegatei, sp. nov., a putative sister species of the extant M. pelagios, based on isolated teeth from late Oligocene–early Miocene (late Chattian–Aquitanian) marine deposits in California and Oregon, U.S.A. Although showing a megachasmid tooth design, teeth of M. applegatei, sp. nov., exhibit a wide morphological range and are reminiscent to those of odontaspidid sharks with strong heterodonty. Megachasma applegatei, sp. nov., could have commonly measured approximately 6 m in total length and likely had a wide range of diet, possibly including small fishes and planktonic invertebrates. The fossil record indicates that either M. applegatei, sp. nov., was broadly adapted to a wide bathymetric tolerance or was a nektopelagic feeder over both deep and shallow water habitats. |
|
New Paper
Recent Papers:
AFONSO, A.S. & HAZIN, F.H.V. (2014): Post-release survival and behavior and exposure to fisheries in juvenile tiger sharks, Galeocerdo cuvier, from the South Atlantic. Journal of Experimental Marine Biology and Ecology, 454: 55-62 http://dx.doi.org/10.1016/j.jembe.2014.02.008
AINSLEY, S.M. & EBERT, D.A. & NATANSON, L.J. & CAILLIET, G.M. (2014): A comparison of age and growth of the Bering skate, Bathyraja interrupta ( Gill and Townsend, 1897), from two Alaskan large marine ecosystems. Fisheries Research, 154: 17-25 http://dx.doi.org/10.1016/j.fishres.2014.02.002
AKHILESH, K.V. & BINEESH, K.K. & MISHRA, S.S. & GANGA, U. & PILLAI, N.G.K. (2014): Notes on the Indian swellshark, Cephaloscyllium silasi (Scyliorhinidae: Carcharhiniformes) from deep waters off the west coast of India. Marine Biodiversity Records, 7: e25 http://dx.doi.org/10.1017/S1755267214000141
ANDREEVA, A.M. (2013): Identification of some proteins of blood and tissue fluid in the fish with enciphered genome. Journal of Evolutionary Biochemistry and Physiology, 49 (6): 551-561 http://dx.doi.org/10.1134/S0022093013060029
BARBIERI, M. & MALTAGLIATI, F. & ROLDÁN, M.I. & CASTELLI, A. (2014): Molecular contribution to stock identification in the small-spotted catshark, Scyliorhinus canicula (Chondrichthyes, Scyliorhinidae). Fisheries Research, 154: 11-16 http://dx.doi.org/10.1016/j.fishres.2014.01.021
BEN BACHA, A. & ABID, I. & HORCHANI, H. & MEJDOUB, H. (2013): Enzymatic properties of stingray Dasyatis pastinaca group V, IIA and IB phospholipases A2: A comparative study. International Journal of Biological Macromolecules, 62: 537-542 http://dx.doi.org/10.1016/j.ijbiomac.2013.10.003
BORNATOWSKI, H. & NAVIA, A.F. & BRAGA, R.R. & ABILHOA, V. & CORRÊA, M.F.M. (2014): Ecological importance of sharks and rays in a structural foodweb analysis in southern Brazil. ICES Journal of Marine Science, in press http://dx.doi.org/10.1093/icesjms/fsu025
BRADLEY, D. & GAINES, S.D. (2014): Counting the cost of overfishing on sharks and rays: Over half of all shark and ray species are at risk of extinction or at least heading that way. eLife, 3: 02199 http://dx.doi.org/10.7554/eLife.02199
BRAUN, C.D. & SKOMAL, G.B. & THORROLD, S.R. & BERUMEN, M.L. (2014): Diving Behavior of the Reef Manta Ray Links Coral Reefs with Adjacent Deep Pelagic Habitats. PLoS ONE, 9 (2): e88170 http://dx.doi.org/10.1371/journal.pone.0088170
CAPAPÉ, C. & MNASRI-SIOUDI, N. & EL KAMEL-MOUTALIBI, O. & BOUMAÏZA, M. & BEN AMOR, M.M. & REYNAUD, C. (2014): Production, maturity, reproductive cycle and fecundity of small-spotted catshark, Scyliorhinus canicula (Chondrichthyes: Scyliorhinidae) from the northern coast of Tunisia (Central Mediterranean). Journal of Ichthyology, 54 (1): 111-126 http://dx.doi.org/10.1134/S0032945214010020
CHEN, X. & AI, W. & XIANG, D. & CHEN, S. (2014): Complete mitochondrial genome of the red stingray Dasyatis akajei (Myliobatiformes: Dasyatidae).Mitochondrial DNA, 25 (1): 37-38 http://dx.doi.org/10.3109/19401736.2013.779262
CLAES, J.M. & NILSSON, D.-E. & STRAUBE, N. & COLLIN, S.P. & MALLEFET, J. (2014): Iso-luminance counterillumination drove bioluminescent shark radiation.Scientific Reports, 4: 4328 http://dx.doi.org/10.1038/srep04328
DE MADDALENA, A. (2013): Great white sharks of South Africa. New Voices Publishing & Alessandro De Maddalena, Western Cape, South Africa, ISBN: 978-0-620-56945-3, 96 pp.
DELPIANI, G.E. & SPATH, M.C. & FIGUEROA, D.E. (2013): Feeding ecology of the southern thorny skate, Amblyraja doellojuradoi on the Argentine Continental Shelf.Journal of the Marine Biological Association of the United Kingdom, 93 (8): 2207-2216 http://dx.doi.org/10.1017/S0025315413000787
DHANEESH, K.V. & ZACHARIA, P.U. (2013): Shark Finning: Are Indian Waters Becoming a Graveyard for Sharks? Journal of Indian Ocean Studies, 21 (3): 358-374
DUMAN, O.V. & BASUSTA, N. (2013): Age and Growth Characteristics of Marbled Electric Ray Torpedo marmorata (Risso, 1810) Inhabiting Iskenderun Bay, North-eastern Mediterranean Sea. Turkish Journal of Fisheries and Aquatic Sciences, 13 (3): 541-549 http://dx.doi.org/10.4194/1303-2712-v13_3_19
ERGÜDEN, D. & GÜRLEK, M. & TURAN, C. (2013): New Mediterranean Marine biodiversity records (December, 2013): 2.8. A young Isurus oxyrinchus Rafinesque, 1810 (Chondrichthyes: Lamnidae) individual captured from Iskenderun Bay, Turkey.Mediterranean Marine Science, 14 (2): 468-469 http://dx.doi.org/10.12681/mms.676
FUJINAMI, Y. & TANAKA, S. (2013): Age, growth and reproduction of the banded houndshark Triakis scyllium around the tip of the Izu Peninsula, Japan. Nippon Suisan Gakkaishi, 79 (6): 968-976
GARNER, M.M. (2013): A Retrospective Study of Disease in Elasmobranchs.Veterinary Pathology, 50 (3): 377-389 http://dx.doi.org/10.1177/0300985813482147
KETCHUM, J.T. & HEARN, A. & KLIMLEY, A.P. & PEÑAHERRERA, C. & ESPINOZA, E. & BESSUDO, S. & SOLER, G. & ARAUZ, R. (2014): Inter-island movements of scalloped hammerhead sharks (Sphyrna lewini) and seasonal connectivity in a marine protected area of the eastern tropical Pacific. Marine Biology, in press http://dx.doi.org/10.1007/s00227-014-2393-y
KHONSARI, R.H. & SEPPALA, M. & PRADEL, A. & DUTEL, H. & CLÉMENT, G. & LEBEDEV, O. & GHAFOOR, S. & ROTHOVA, M. & TUCKER, A. & MAISEY, J.G. & FAN, C.-M. & KAWASAKI, M. & OHAZAMA, A. & TAFFOREAU, P. & FRANCO, B. & HELMS, J. & HAYCRAFT, C.J. & DAVID, A. & JANVIER, P. & COBOURNE, M.T. & SHARPE, P.T. (2013): The buccohypophyseal canal is an ancestral vertebrate trait maintained by modulation in sonic hedgehog signaling. BMC Biology, 11: 27 http://dx.doi.org/10.1186/1741-7007-11-27
LESNIAK, T.C. & SCHIRMER, A.E. & KNAPP, C.R. (2013): Ultrasound evidence of fetal resorption in the yellow stingray, Urobatis jamaicensis (Urotrygonidae). Cybium, 37 (4): 314-316
LEVEELAHTI, L. & RYTKONEN, K.T. & RENSHAW, G.M.C. & NIKINMAA, M. (2014): Revisiting redox-active antioxidant defenses in response to hypoxic challenge in both hypoxia-tolerant and hypoxia-sensitive fish species. Fish Physiology and Biochemistry, 40 (1): 183-191 http://dx.doi.org/10.1007/s10695-013-9835-1
LOBODA, T.S. & DE CARVALHO, M.R. (2013): Systematic revision of the Potamotrygon motoro (Müller & Henle, 1841) species complex in the Paraná-Paraguay basin, with description of two new ocellated species (Chondrichthyes: Myliobatiformes: Potamotrygonidae). Neotropical Ichthyology, 11 (4): 693-737
NAWAZ, S. & SCHWEITZER, J. & JAHN, O. & WERNER, H.B. (2013): Molecular evolution of myelin basic protein, an abundant structural myelincomponent. Glia, 61 (8): 1364-1377
NOSAL, A.P. & CAILLAT, A. & KISFALUDY, E.K. & ROYER, M.A. & WEGNER, N.C. (2014): Aggregation behavior and seasonal philopatry in male and female leopard sharks Triakis semifasciata along the open coast of southern California, USA. Marine Ecology Progress Series, 499: 157-175 http://dx.doi.org/10.3354/meps10632
PRETORIUS, C. & GRIFFITHS, C.L. (2013): Patterns of egg deposition and egg development in the catsharks Poroderma pantherinum and Haploblepharus pictus.African Zoology, 48 (1): 115-124
RÊGO, M.G. & HAZIN, F.H.V. & NETO, J.E. & OLIVEIRA, P.G.V. & SOARES, M.G. & TORRES, K.R.L.S. & LANA, F.O. & ROQUE, P.C.G. & SANTOS, N.L. & COELHO, R. (2013): Morphological analysis and description of the ovaries of female silky sharks, Carcharhinus falciformis (Müller & Henle, 1839). Neotropical Ichthyology, 11 (4): 815-819 http://dx.doi.org/10.1590/S1679-62252013000400009
SCHAFFER, P.A. & LIFLAND, B. & VAN SOMMERAN, S. & CASPER, D.R. & DAVIS, C.R. (2013): Meningoencephalitis Associated With Carnobacterium maltaromaticum–Like Bacteria in Stranded Juvenile Salmon Sharks (Lamna ditropis).Veterinary Pathology, 50 (3): 412-417 http://dx.doi.org/10.1177/0300985812441033
SPERONE, E. & COPPOLA, F. & GIGLIO, G. & CIRCOSTA, V. & MICARELLI, P. & TRIPEPI, S. & COMPAGNO, L.J.V. (2013): New Mediterranean Marine biodiversity records (December, 2013): 2.6. On the occurrence of the dusky shark Carcharhinus obscurus in Calabria (Central Mediterrane an, Southern Italy). Mediterranean Marine Science, 14 (2): 467-468 http://dx.doi.org/10.12681/mms.676
TSIKLIRAS, A.C. & STERGIOU, K.I. (2014): Size at maturity of Mediterranean marine fishes. Reviews in Fish Biology and Fisheries, 24 (1): 219-268 http://dx.doi.org/10.1007/s11160-013-9330-x
VANDEPERRE, F. & AIRES-DA-SILVA, A. & SANTOS, M. & FERREIRA, R. & BOLTEN, A.B. & SANTOS, R.S. & AFONSO, P. (2014): Demography and ecology of blue shark (Prionace glauca) in the central North Atlantic. Fisheries Research, 153: 89-102 http://dx.doi.org/10.1016/j.fishres.2014.01.006
VAUGHAN, N.H. & SMITH, S.L. (2013): Isolation and characterization of a c-type lysozyme from the nurse shark. Fish & Shellfish Immunology, 35 (6): 1824-1828 http://dx.doi.org/10.1016/j.fsi.2013.09.001
WANG, Y. & ZHANG, M. & WANG, C. & YE, B. & HUA, Z. (2013): Molecular cloning of the alpha subunit of complement component C8 (CpC8α) of whitespotted bamboo shark (Chiloscyllium plagiosum). Fish & Shellfish Immunology, 35 (6): 1993-2000 http://dx.doi.org/10.1016/j.fsi.2013.09.010
WARD-PAIGE, C.A. & BRITTEN, G.L. & BETHEA, D.M. & CARLSON, J.K. (2014): Characterizing and predicting essential habitat features for juvenile coastal sharks.Marine Ecology, in press http://dx.doi.org/10.1111/maec.12151
WEIGMANN, S. & STEHMANN, M. & THIEL, R. (2014): Contribution to the taxonomy and distribution of Pristiophorus nancyae (Elasmobranchii: Pristiophoriformes) from the deep western Indian Ocean. Marine Biodiversity, in press http://dx.doi.org/10.1007/s12526-013-0200-5
WEIGMANN, S. & STEHMANN, M. & THIEL, R. (2014): Complementary redescription of Anacanthobatis ori (Wallace, 1967) and its assignment to Indobatis n. g. (Elasmobranchii, Anacanthobatidae), with comments on other legskates. Zootaxa, 3779 (2): 101-132 http://dx.doi.org/10.11646/zootaxa.3779.2.1
Parasitology:
BAKOPOULOS, V. & KSIDIA, V.-C. (2014): Pontobdella muricata infection of Raja clavata and Dasyatis pastinaca off the coast of Lesvos, Greece. Journal of the Marine Biological Association of the United Kingdom, 94 (2): 405-409 http://dx.doi.org/10.1017/S0025315413000830
CAIRA, J.N. & JENSEN, K. & WAESCHENBACH, A. & OLSON, P.D. & LITTLEWOOD, D.T.J. (2014): Orders out of chaos - molecular phylogenetics reveals the complexity of shark and stingray tapeworm relationships. International Journal for Parasitology, 44 (1): 55-73 http://dx.doi.org/10.1016/j.ijpara.2013.10.004
CAIRA, J.N. & PICKERING, M. (2013): Cestodes from deep-water squaliform sharks in the Azores. Deep Sea Research Part II: Topical Studies in Oceanography, 98: 170-177 http://dx.doi.org/10.1016/j.dsr2.2013.08.008
Fossil:
BRAZEAU, M.D. & FRIEDMAN, M. (2014): The characters of Palaeozoic jawed vertebrates. Zoological Journal of the Linnean Society, in press http://dx.doi.org/10.1111/zoj.12111
FERRÓN, H. & PLA, C. & MARTÍNEZ-PÉREZ, C. & ESCUDERO-MOZO, M.J. & BOTELLA, H. (2014): Morphometric Discriminant Analysis of isolated chondrichthyan scales for palaeoecological inferences: the Middle Triassic of the Iberian Chain (Spain) as a case of study. Journal of Iberian Geology, 40 (1): 87-97 http://dx.doi.org/10.5209/rev_JIGE.2014.v40.n1.44089
HERMAN, J. & VAN WAES, H. (2014): Observations concerning the Evolution and the Parasystematics of all the living and fossil Chlamydoselachiformes, Squatiniformes, Orectolobiformes, and Pristiophoriformes, based on both biological and odontological data. Suggestion of a possible origin of the Order Pristiophoriformes, of the Order Ganopristiformes and a global Synthesis of the previous Systematics proposals. Géominpal Belgica, 6: 347 pp
KHONSARI, R.H. & SEPPALA, M. & PRADEL, A. & DUTEL, H. & CLÉMENT, G. & LEBEDEV, O. & GHAFOOR, S. & ROTHOVA, M. & TUCKER, A. & MAISEY, J.G. & FAN, C.-M. & KAWASAKI, M. & OHAZAMA, A. & TAFFOREAU, P. & FRANCO, B. & HELMS, J. & HAYCRAFT, C.J. & DAVID, A. & JANVIER, P. & COBOURNE, M.T. & SHARPE, P.T. (2013): The buccohypophyseal canal is an ancestral vertebrate trait maintained by modulation in sonic hedgehog signaling. BMC Biology, 11: 27 http://dx.doi.org/10.1186/1741-7007-11-27
MAISEY, J.G. (2014): The diversity of tessellated calcification in modern and extinct chondrichthyans. Revues de Paleobiologie, in press
MAISEY, J.G. & TURNER, S. & NAYLOR, G.J.P. & MILLER, R. (2014): Dental patterning in the earliest sharks: implications for tooth evolution. Journal of Morphology, in press http://dx.doi.org/10.1002/jmor.20242
PRADEL, A. & MAISEY, J.G. & TAFFOREAU, P. & MAPES, R.H. & MALLATT, J. (2014): A Palaeozoic shark with osteichthyan-like branchial arches. Nature (In press).
PŘIKRYL, T. (2013): An annotated list of the Oligocene fish fauna from the Osíčko locality (Menilitic Fm.; Moravia, the Czech Republic). Acta Musei Nationalis Pragae, Series B - Historia Naturalis, 69 (3-4): 195-203 http://dx.doi.org/10.14446/AMNP.2013.195
SHIMADA, K. & WELTON, B.J. & LONG, D.J. (2014): A new fossil megamouth shark (Lamniformes, Megachasmidae) from the Oligocene-Miocene of the western United States. Journal of Vertebrate Paleontology, 34 (2): 281-290 http://dx.doi.org/10.1080/02724634.2013.803975
SOLER-GIJÓN, R. & POYATO-ARIZA, F.J. & MAISEY, J.G. & LANE, J.A. (2014): Chondrichthyes. In: Poyato-Ariza, F.J., Buscalioni, A.D. & Fregenal-Martinez, M. (eds), Las Hoyas: A Cretaceous wetland. A multidisciplinary synthesis after 25 years of research on an exceptional fossil deposit from Spain, in press
|
|
source: http://www.irishexaminer.com/world/big-leap-as-australia-hatches-first-ivf-shark-261828.html
‘Big leap’ as Australia hatches first IVF shark
Thursday, March 13, 2014
Australia has successfully hatched its first shark born via artificial insemination with hopes that the development can ultimately be used to help breed threatened species, an aquarium said.
Sea Life Melbourne Aquarium said the brown banded bamboo shark pup was born on March 3, ending a process which began in September when aquarists collected a semen sample from a shark in Mooloolaba in northeastern Australia.
This was flown to the southern city of Melbourne and inseminated into the mother the same day — making the pup the first shark to be born globally via a live semen sample transported from one facility to another, Sea Life said.
Melbourne Aquarium vet Rob Jones said the birth of the shark — which is expected to grow from its initial length of 16cm (6.3in) to an adult size of 1.2 to 1.5m — was a milestone. “This is a big leap,” he said.
The hatching is part of a nine-year project into understanding the reproductive behaviours of sharks, animals which are common in Australia but are little understood.
The team hope their research will help with plans to manage threatened species in the wild, in particular the critically endangered grey nurse shark.
© Irish Examiner Ltd. All rights reserved
|
|
source: http://www.sciencedaily.com/releases/2014/03/140304095107.htm
Fish bioluminescence: Distinctive flashing patterns might facilitate fish mating
This is a blue lanternfish (Tarletonbeania crenularis), which is found in the Eastern Pacific. The photophores, light-producing structures, can be seen as spots on the side and belly of the fish.
Credit: Copyright Matthew P. Davi
Scientists have shown for the first time that deep-sea fishes that use bioluminescence for communication are diversifying into different species faster than other glowing fishes that use light for camouflage. The new research indicates that bioluminescence -- a phenomenon in which animals generate visible light through a chemical reaction -- could promote communication and mating in the open ocean, an environment with few barriers to reproduction. The study was recently published in the journalMarine Biology.
"Bioluminescence is quite common in the deep sea, and many fishes inhabiting this region exhibit complex, species-specific patterns of light-producing structures," said John Sparks, a curator in the American Museum of Natural History's Department of Ichthyology and one of the co-authors on the study. "But we still have so much to learn about how these animals use bioluminescence -- for predation, camouflage, communication, or something else. This new work provides insight into how this phenomenon might have shaped present-day biodiversity in the deep open ocean."
Unlike on land, where rivers, mountain ranges, and other physical obstacles can genetically isolate animals from one another resulting in speciation events over time, in the deep open ocean there are few obvious physical barriers to reproduction and gene flow. This has traditionally been considered one of the reasons why there is a comparatively low level of fish species richness in the deep sea. For example, bristlemouths, which are among the most abundant vertebrates on Earth, are represented by only 21 species. But that's not the case for all fishes. Lanternfishes, which inhabit the same mid-water, or mesopelagic, area of the ocean, have diversified into more than 250 species.
"The comparison of lanternfishes and bristlemouths is ideal for studying speciation in the deep sea. Both bioluminescent groups are among the most abundant vertebrates on Earth and live in the same dark environment," said Matthew Davis, a research associate at the University of Kansas and the study's lead author. "The difference in species numbers between these two groups is striking. Both use bioluminescence for camouflage, but lanternfishes have evolved a suite of light organs that act as a beacon for communication, which our work suggests have had an incredible impact on their diversification in the deep sea."
To investigate, Sparks, Davis, and other scientists from the University of Kansas and Johnson County Community College reconstructed a tree of life for ray-finned fishes with a particular focus on the evolution of bioluminescence.
Many fishes emit light from organs called photophores that appear as luminous spots on the body. On lanternfishes, photophores are present ventrally along the belly, laterally on the flank and head, and on the tail. The researchers discovered that the common ancestor of lanternfishes most likely evolved this complex photophore system during the Late Cretaceous, between 73-104 million years ago.
The significance of the photophores on the underside of mesopelagic fishes has long been thought to provide camouflage against predators swimming below, helping them to blend in with any residual light shining down from the surface. But the function of photophores on the side of the body has been obscure, until now. Using mathematical techniques based on the anatomy of the fishes, the researchers determined that the lateral photophore patterns on certain lanternfish lineages are distinct enough to allow identification of individual species. This is not the case for photophores on the belly. Recent work has shown that lanternfishes are capable of seeing blue-green bioluminescence from up to about 100 feet away, supporting the idea that lateral photophores could be used for interspecific communication.
"In this study we have shown that deep-sea fishes that exhibit unique, species-specific bioluminescent organs, like lanternfishes and dragonfishes, also exhibit increased rates of diversification," said Leo Smith, an assistant curator of ichthyology at the University of Kansas and a co-author on the paper. "This suggests to us that bioluminescent signaling may be critical to diversification of fishes in the deep sea."
To further test this hypothesis, the researchers plan to record lanternfish flashing patterns using emerging technology, such as remotely operated vehicles outfitted with ultra low-light underwater cameras. Other tools that might assist in this type of research include the Exosuit, a next-generation, human-piloted atmospheric diving system now on display in the American Museum of Natural History's Irma and Paul Milstein Family Hall of Ocean Life through March 5, 2014.
Story Source:
The above story is based on materials provided by American Museum of Natural History. Note: Materials may be edited for content and length.
Journal Reference:
- Matthew P. Davis, Nancy I. Holcroft, Edward O. Wiley, John S. Sparks, W. Leo Smith. Species-specific bioluminescence facilitates speciation in the deep sea. Marine Biology, 2014; DOI: 10.1007/s00227-014-2406-x
|
|
source: http://www.sciencedaily.com/releases/2014/02/140227142615.htm
'Shark's eye' view: Witnessing the life of a top predator
Credit: Mark Royer/University of Hawaii
Instruments strapped onto and ingested by sharks are revealing novel insights into how one of the most feared and least understood ocean predators swims, eats and lives.
For the first time, researchers at the University of Hawaii and the University of Tokyo outfitted sharks with sophisticated sensors and video recorders to measure and see where they are going, how they are getting there, and what they are doing once they reach their destinations.
Scientists are also piloting a project using instruments ingested by sharks and other top ocean predators, like tuna, to gain new awareness into these animals' feeding habits. The instruments, which use electrical measurements to track ingestion and digestion of prey, can help researchers understand where, when and how much sharks and other predators are eating, and what they are feasting on.
The instruments are providing scientists with a "shark's eye" view of the ocean and greater understanding than ever before of the lives of these fish in their natural environment.
"What we are doing is really trying to fill out the detail of what their role is in the ocean," said Carl Meyer, an assistant researcher at the Hawaii Institute of Marine Biology at the University of Hawaii at Manoa. "It is all about getting a much deeper understanding of sharks' ecological role in the ocean, which is important to the health of the ocean and, by extension, to our own well-being."
Using the sensors and video recorders, the researchers captured unprecedented images of sharks of different species swimming in schools, interacting with other fish and moving in repetitive loops across the sea bed. They also found that sharks used powered swimming more often than a gliding motion to move through the ocean, contrary to what scientists had previously thought, and that deep-sea sharks swim in slow motion compared to shallow water species.
"These instrument packages are like flight data recorders for sharks," Meyer said. "They allow us to quantify a variety of different things that we haven't been able to quantify before."
"It has really drawn back the veil on what these animals do and answered some longstanding questions," he added.
Meyer and Kim Holland, a researcher also at the Hawaii Institute of Marine Biology, are presenting the new research today at the 2014 Ocean Sciences Meeting co-sponsored by the Association for the Sciences of Limnology and Oceanography, The Oceanography Society and the American Geophysical Union.
Sharks are at the top of the ocean food chain, Meyer noted, making them an important part of the marine ecosystem, and knowing more about these fish helps scientists better understand the flow of energy through the ocean. Until now, sharks have mainly been observed in captivity, and have been tracked only to see where they traveled.
These new observations could help shape conservation and resource management efforts, and inform public safety measures, Holland said. The instruments being used by scientists to study feeding habits could also have commercial uses, including for aquaculture, he added.
Story Source:
The above story is based on materials provided by American Geophysical Union.Note: Materials may be edited for content and length.
|
|
|